There are 3 probable adult males (NZ.9.3 rear. M1 with associated pupal exuvia, and NZ.17.1 reared M2 and M3):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| Ta5/Ti
|
PI
| 1433
| 1305
| 1823
| 1062
| 787
| 628
| 302
| 1.35-1.38
| 1.06-1.12
| 0.22
|
PII
| 1530
| 1455
| 850
| 577
| 370
| 260
| 190
| 0.56-0.61
| 1.03-1.08
| -
|
PIII
| 1820
| 1870
| 1200
| 770
| 540
| 340
| 210
| 0.62-0.70
| 0.97-5-0.99
| -
|
Abdomen with seg II darkened over anterior half and progressively further posterior on III-V, then whole tergite darkened. 5-10 setae in individual pale areas on TIX.
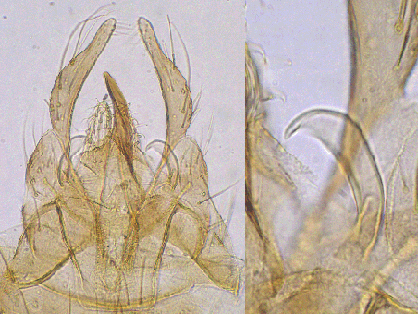
Superior volsella hooked, E(h)-type of Strenzke (1959); Inferior volsella reaching about to the tip of the relatively narrow anal point, but to about basal third of gonostyle, with simple setae. Anal point is quite narrow at base; gonostyle only moderately swollen (width/length 0.19-0.24) and narrows over distal third to half.
Possible pupa: Exuvial length about 8.8 mm; length of inner margin of wing case about 1.8 mm. Indications of small frontal warts. Cephalic tubercles abt 86x132 micron (l/w 0.65).
About 95 recurved hooks on segment II. L-seta on margin of III-IV at least 101 µm. Pedes spurii A of seg. IV about 197x104 µm and about 0.27 of segment length; those of segments V and VI not visible. Caudolateral spur of segment VIII with 6 & 7 appressed spines. About 96 taeniae in incomplete double row, then a double row at posterior end of the hair fringe on each side of the swim fin. Shagreen quite heavy post 1/2 II, 3/4 III then all IV-VI, weak post VI and 2 post patches around seta on VIII; Muscle scars on tergites faint.
Fourth instar larva: A bathophilus-type larva. Length about 17.2-18 mm (female) and 13.8-16.0 (male); smaller larvae from thermal habitats. Anterior ventral tubules about 0.96 (0.61-1.60) mm; posterior about 0.93 (0.25-1.46) mm. Dorsal Anal tubules possibly longer (205-367 µm) than the ventral pair (200-321 µm), both about 1.6-2.2 times longer than wide.
Head capsule with pale or slightly dark posterior third of gula; with pale or slightly darkened; frontoclypeus usually pale but may be slightly darkened.
Mentum (Fig. b) with 4th laterals reduced (type 1-II), generally to height of 5th laterals; central tooth with c2 teeth relatively well separated (type III or IIA if worn).
Ventromental plates (Fig. c) about 191 (182-199) µm wide and 3.5 (3.3-3.65) times wider than deep; 1.05 (0.92-1.17) times the mentum width; IPD about 83.5 (51-101) µm (0.4 of mentum width), with about 41 (36-45) striae. Pecten epipharyngis (Fig. a) with about 12.7 (11-15) often irregular teeth.
Premandible with inner tooth about 4.25-5 times the width of the outer tooth, outer tooth narrowing to a fine point (usually type D, but may be ty. C)
Antenna (Fig. d) with A1 about 0.38 (0.31-0.49) of Ventral head length, and 4.0 (3.3-4.8) times longer than wide; AR about 2.44 (2.33-2.70); Ring Organ about one third to two fifths up from base of segment 1, which is about 4.0 (3.3-4.8) times longer than wide; AR abt 2.44-2.78; ratio of antennal segments (micron): 160 : 35 : 9 : 13 : 7.5.
Mandible (Fig. e) about 292 (230-327) µm long, with 3rd inner tooth partly darkened and only slightly separated (type IIB); about 18 (14-26) grooves on the outer surface near the base, 12 (10-14) striae in the PecM; Mdt-Mat 23-38, MTR 0.31-0.32.
Third instar larva: A single larva was available, anterior Ventral tubules 1.25 mm long. Gula and frontoclypeus pale. Mentum width about 0.6 of Ventral head length.
Ventromental plates by about 0.4 of the mentum width; 132 µm wide; 3.1 times wider than deep, and about the same width as the mentum, with 25-27 striae; VMR 0.35-0.36.
Premandible with inner tooth about 2.5 times wider than the outer tooth. Pecten epipharyngis with 12 teeth.
Distance between antennal bases 100 µm, about 0.95 of that between the S4 setae. The relationships of A1 to the VHL and the distance of the RO from base about the same as in the 4th instar; AR about 1.5, relative length of segments (µm) 83.5 : 32 : 9.5 : 12.5: 6.
Mandible about 190 µm long, of type IIB; with 13 furrows and 10-11 taeniae in the Pecten mandibularis; Mdt-Mat 28 µm, MTR 0.42.
Cytology: 4 polytene chromosomes with the pseudothummi arm combination (AE, BF, CD, G). Arm G without a nucleolus, but with an obvious subterminal BR. Nucleolus developed in arm F at about group 19.
Polymorphism in arm B, with B2 apparently the more common sequence.
Arm A1: 1a-e, 11 - 10, 2c - 1f, 3e - 2e, 7 - 4, 12a-c, 3i-f, 9 - 8, 2d, 13 - 19 as nzlA2
Arm B1: Puff (group 7) with reduced dark bands nearer distal end of arm. as nzl B2
Arm B2: Distal inversion including the puff (group 7) that returns it to near the middle of the arm. as nzlB3
Arm C1: as nzlC3
Arm D1: 1 - 2, 16c-a , 17e-a, 10c-a, 3e - 9, 3d-a, 10d -15, 18 - 24 as nzlD2
Arm E1: 1a-c, 5 - 7c, 10g-c, 3f - 4, 1d - 3e, 10b - 7d, 11 - 13 as nzlE3
Arm F1: 1a-e, 12 - 15c, 2c - 10, 2a, 11i-a, 2b, 15d - 23 as nzlF2
Arm G1: No nucleolus, but an obvious subterminal BR. as nzlG2?
Localities:
North Island:
Lake Ngapouri (-38.00°S, 176.50°E), Waiotapu, South Auckland (NZ.9.1) (D.J. Forsyth) 15-i-1972; D.J. Forsyth & Jon Martin) 5-xii-1973;
and S. Ibarrarán) 29-viii-2007.
Lake Rotoiti (-37.28°S, 174.67°E), about 16 Km north east of Rotorua, South Auckland (NZ.19.1) (Jon Martin, D.J. Forsyth 7-xii-1973;
and S. Ibarrarán) 29-viii-2007.
?Ngapuna (-38.15oS, 176.27oE), Rotorua, South Auckland (NZ.17.1) (Jon Martin,
D.J. Forsyth) 7-xii-1973.
Kinloch Marina, Lake Taupo, South Auckland (NZ.12.1) (D.J.Forsyth) 18-ix-1972
Waikato area (-37.802°S, 175.334°E) (I. Hogg) 16.vii.2012 and (E.Doyle & N. Binks) 12.xi.2013.
South Island:
Lake Rotoiti (-41.82°S, 172.84°E), Nelson Lakes National Park, Tasman region
(NZ.83.1) (B.V. Timms) 1-x-1968.
Initially included with C. novae-zelandiae, but shows consistent differences in the lighter larval head colour and some unique banding patterns. The existence of this species was partially supported by the initial barcoding analyses of Sofia Ibarrarán, and there are some differences in the ITS-1 sequence. More detailed analysis of the CO1 barcode sequence places it in the C. novaezelandiae complex (see under C. novaezelandiae) but differing at a number of base positions. The existence of polymorphic sites in C. novaezelandiae may result in some specimens being incorrectly assigned to the C.spNZ12 Bin.
See also C. novaezelandiae, C. species NZ10, and C. 'spilleri'.[ Return to Key| Go to References ]