9. Chironomus n.sp. NZ9 (C. 'rotorua')In BOLD Bin: BOLD:AAL7010
The nearest neighbor BIN is listed as BOLD: AAL7011, but this Bin does not seem to exist any longer and the replacement Bin (BOLD:ACK7128) also does not exist.Adult: Male not known. In the absence of associated males, the potential Holotype is a larva with polytene chromosomes from Lake Hayes, Queenstown, NZ.47.1 Sl. 21F. Female: (from a specimen collected by I. Hogg, in BOLD database): Wing length about 4.3 mm., anterior wing veins darkened. Colour brown with darker brown postnotum and dark bands across basal part of abdominal segments at least II-VI.
Legs apparently relatively pale with no darkening at knees. Mid femur abt. 1480 µm; mid tibia about 1240 µm (F/T abt 1.19); hind femur abt. 1920 µm, tibia at least 1900 µm (F/T about 1). Pupa not known Fourth instar larva: salinarius type larva, length 15.5-15.8 mm (female), 12.5-15.7 mm (male). Head capsule with gular region darkened on posterior third, frontoclypeus very slightly to slightly darkened, but with some darkening of the posterior border of the frontoclypeus.
Anal tubules about 300-360 µm long, without a constriction, and about 1.5-2.3 times longer than wide, dorsal and ventral pairs generally similar in size. Oesophageal opening 83-124 µm wide and 2.9-4.9 times wider than deep.
Mentum (Fig. d, below) with c1 tooth characteristically broad and flat on top, c2 teeth relatively well separated (type broad IIA); 4th lateral reduced to about the level of the 5th lateral (type II), 6th lateral arising at a slightly lower level than other teeth; width 0.71-0.77 of VHL (i.e. head relatively wide).
Ventromentum (Fig. e, below) about 3.6 times wider than deep; with about 36-40 striae; VMR about 0.35-0.38. Pecten epipharyngis (Fig. a, below) with about 14-15 relatively even teeth (Ty. B). Premandible (Fig. b, below) with robust teeth, inner tooth about 4-5 times wider than the outer tooth, which narrows to a fine point when not worn.
Antenna (Fig. c, below) with basal segment about 0.4 of the VHL; about 2.81 (2.6-3.2) x longer than wide, RO about a third (0.29-0.36) up from base; AR 1.84 (1.59-2.07); antennal proportions (micron) 125 : 35 : 10 : 15 : 7.
Distance between the antennal bases (199.75 µm) generally similar to that between the S4 setae (204.88 µm) but quite variable; S4 setae separated by 87-95% of the frontoclypeus width, i.e. often very close to the edge of the frontoclypeus. S5 setae about level, or anterior to nearby RO.
Mandible (Fig. f, below) about 300 (245-328) µm in length; 3rd inner tooth only partly separated and coloured (type IIB), with about 22.5 (18-26) furrows on outer surface near the base; Pecten mandibularis of 12.4 (11-14) taeniae; Mdt-Mat 22.5-40.5 µm, MTR about 0.26-0.36, dependant on degree of tooth wear. 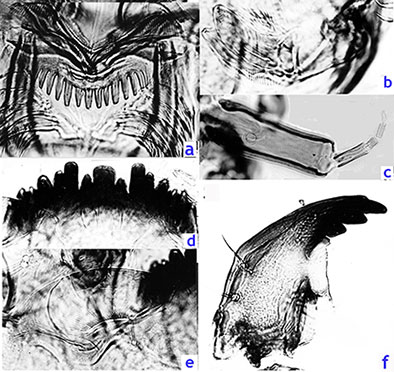 Gut of most larvae almost filled with sand particles. Cytology: 4 polytene chromosomes with thummi cytocomplex arm combination (AB, CD, EF, G).
Centromeres heterochromatic. Two nucleoli, one proximal in arm F; the other about one third from the heterochromatic end of arm G. A Balbiani ring developed near distal end of arm G. Arm A with the sequence A4 found in Australian species; arm E with sequence E1 of the Australian species; and arm F as in C. oppositus F3 and C. australis.. No polymorphism in the small sample examined. Arm A1: 1a-e, 11-10, 2c-1f, 3e-2d, 8-9, 3f-i, 12c-a, 4-7, 13-19 ie. as A4 oppositus
Arm B1: Puff (group 7) and about 2 distal dark bands (8ab) , close to 4 characteristic bands.
Arm C1: possibly 10A-10D6, 11B11-10D7, 11B12-11E; ie. as C2 of forsythi
Arm D1: 1 - 2, 16c-a,17e-a, 10c-a, 3e - 9, 3d-a, 10d -15, 18 - 24 ie. as forD1, nzlD2
Arm E1: 1 - 3e, 10b - 3f, 10c - 13 ie. as E1 oppositus, analis, #&39;castaneum'-gp., etc.
Arm F1: 1 - 2a, 10 - 2c, 15c - 11a, 2b, 15d - 23 ie. as F3 of oppositus F3, australis, analis, forsythi, 'castaneum'-gp., etc.
Arm G1: heterochromatic terminal centromere, small nucleolus (or BR?) in middle and BR near the distal end Localities:
North Island:
Lake Okaro (38.30°S, 176.40°E), Rotorua area (NZ.10.8 & NZ.10.9) (D.J.Forsyth & S. Ibarrarán) 14-ix-1982 and 14-ii-2007
Lake Okareka (38.285°S, 176.603°E), Rotorua area (S. Ibarraran) 15-ii-2007
n. Te Awamutu, Waikato area (37.802°S, 175.334°E) (Ian Hogg) 29-iv-2013South Island:
Lake Hayes (44.98°S, 168.80°E), c.13 km n.e. Queenstown, Westland (NZ.47.1) (Jon Martin) 7-i-1974MtCOI sequence suggests that there may be hybridization with C. 'castaneum' particularly where they occur together in the same lake, although this is unlikely due to the whole arm translocation difference. On the other hand, there is a possibility that this is a recent whole-arm translocation event from a related pseudothummi-complex species (probably C. 'castaneum', sp. 6a, on the basis of current information – see below), with no connection to species of the thummi-cytocomplex. As such, this species would be immediately reproductively isolated from its pseudothummi-cytocomplex parent. The limited mitochondrial DNA difference would then reflect that this event occurred relatively recently. There are differences at 14 base positions between the three members of the 'castaneum-group': Base
C. 'rotorua' (spNZ9)
G GC A A T G T T A C/G T G A T
C. 'castaneum' (spNZ6a)
A GC A/G A/G T/A G T/C T/C A/T/G G/A T/A G/A A/G T/G
C. 'paracastaneum' (spNZ6b)
A AA G G A A C C G A A A G G The similarity of many of the polymorphic bases of C. 'castaneum' with the equivalent base of spNZ9 suggests that it is the ancestral species in which the translocation occurred. On the other hand, the similarity of the polymorphic bases to those of C. 'paracastaneum' could be the result of hybridization between these two closely related species. [ Go to C. 'castaneum' | Return to Key| Go to References ] |