5. Chironomus 'thermarum' Manuscript name by D. J. Forsyth.In Bold Bin: BOLD:AAJ0168 or BOLD:ABZ5458This is a morphologically variable species in regards to larval type. Adult:
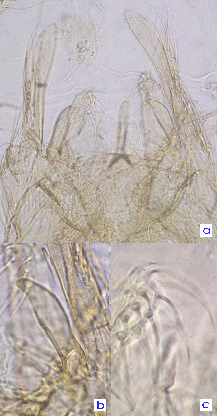
Forsyth describes the abdominal tergites as largely dark with only a narrow pale posterior margin of segments II to VI and on four fifths of segments VII and VIII. A pupa with a pharate male is available, from which the following characters could be determined: Tergite IX with 5 setae in individual pale areas. Anal point (a, above) narrow at base, but appears turned down at the end which is broader; Superior volsella (b, above) with just a small hook at the tip (D(e)-type of Strenzke 1959); Inferior volsella (a, above) extending to about middle of gonostylus with setae simple (c, above). Gonostylus (a, above) moderately broad but narrowing only slightly at distal end, about 4+1 setae at tip. Pupa: about 6.9 mm (1 male)-8.3 mm (1 female) long; inner margin of wing case about 1.44-1.66 mm. Cephalic tubercle about 85-114 µm long. Basal ring about 115-145 µm long and 57-65 µm wide, HR 1.77-2.56. Thorax and muscle scars yellow-brown, otherwise pale.
Shagreen on posterior half of segment II, increasing to whole of segments IV-VI, little on segments VII & VIII. Hook row with 70-92 hooks, occupying about 0.6-0.8 of segment width. PSB on segs II and III; PSA of segment IV large (202-218 µm long, 101-145 µm wide) and about 0.26-0.28 length of segment. Caudolateral spur of segment VIII (below) with about 2 or 3 spines, at least 1 small. 68-93 taeniae on each side of anal lobe, mostly in a single row, but some double at posterior end. Fourth instar larva: halophilus-type, i.e. with some development of posterior pair of ventral tubules (length about 120 µm), or sometimes salinarius-type. However some specimens consistent with C. 'thermarum' have been found that are essentially bathophilus-type, with both short anterior and posterior VT (over 0.5 mm in length). Smaller species (temperature effect?), length about 11.3 mm (F), 9.2 mm (M)). Anal tubules from 1.5-2.1 times longer than wide.
Head capsule with frontoclypeus generally pale or slightly darkened, gula generallydarkened.
Mentum (Fig. b) usually with c2 teeth only partly separated (type IB); ; 4th laterals only slightly reduced (type II); 6th laterals arising from a lower level Mentum width about half VHL.
Ventromentum (below) with about 45-55 striae; VMR 0.29-0.32. Pecten epipharyngis with about 11-14 irregular teeth. Basal antennal segment about 3.27-3.68 times as long as wide; AR 2.48 (2.22-2.66); segment proportions (micron): 124 : 23 : 8 : 12 : 7.
Distance between antennal bases less than that between the S4 setae
Mandible (Fig. e) about 220-252 µm long (heel to Mdt) of type II, with about 13-15 furrows on outer surface near base; 11-13 taeniae in Pecten mandibularis; Mdt-Mat 22-30 µm; MTR 0.30-0.43. Cytology: 4 polytene chromosomes, pseudothummi-complex arm combination (BF,CD,EA,G).
Arm G normally closely paired with a subterminal nucleolus; another large nucleolus on arm F. Only one Balbiani ring (BR), in the middle of arm G, was visible in the specimens examined.
Arm A with sequence A4 as found in Australian species; arm E with sequence E1 of Australian species, and arm C appears to have the same sequence as C1 of C. forsythi (ie. as C1 of the Australian species).
No polymorphism was seen in the available specimens.thmA1: 1a-e, 11-10, 2c-1f, 3e-2d, 8-9, 3f-i, 12c-a, 4-7, 13-19 ie. as A4 of oppositus, nzlA1, AnlA1, forA1
thmB1: Puff (group 7) hardly developed, with proximal dark bands (group 8) near distal end. as nzlB4
thmB2: possibly as oppositus BI. possibly in non-thermal habitats
thmC1: as C1 of forsythi and oppositus?
thmD1: 1 - 2, 16c-a,17e-a, 10c-a, 3e - 9, 3d-a, 10d -15, 18 - 24
thmE1: 1 - 3e, 10b - 3f, 10c - 13 ie. as oppositus E1
thmF1: appears to be 1 - 2a, 10 - 2c, 15c - 11a, 2b, 15d - 23 ie. as oppositus E1
thmG1: Subterminal nucleolus and a median BR. Molecular:Mt COI: Sequences suggest that C. 'thermarium' may be an ecologically plastic species that can live in thermal habitats as well as habitats with lower water temperatures. At normal temperatures the larvae are a bathophilus type, with the ventral tubules, particularly the anterior pair, reducing in size with higher temperatures (see below). Localities:
North Island:
Kerosene Creek (thermal), South Auckland (38.94°S, 176.56°E) (NZ.78.1) (S.Ibarraran) 27-viii-2007
Lake Ngahewa (38.31°S, 176.37°E), South Auckland (NZ.76.1) (S. Ibarraran) 28-viii-2007.
Lake Rotowhero, South Auckland (39.30°S, 176.40°E) (NZ.11.3) (D.J. Forsyth and Jon Martin) 23-xii-1973; (NZ.11.6) (S. Ibarraran) abt.15-ii-2007
Queen Elizabeth Park (37.94°S,175.56°E), South Auckland (NZ.80.1) (S. Ibarraran) abt 15-ii-2007. Waiotapu Stream (thermal), South Auckland (38.38°S, 176.35°E) (NZ.69.1) (D.J.Forsyth) 14-vi-1983
Waiotapu Stream (thermal), South Auckland (38.38°S, 176.35°E) (NZ.69.2) (D.J.Forsyth) 25-iii-1998 [ Go to C. novaezelandiae | Return to Main File| Go to References ] |