Synonyms: C. crassimanus Keyl 1962.In BOLD Bin: BOLD:AAJ4234.
Specimens from North America are placed in a separate Bin (see below)Adult
Adults of Japanese specimens are entirely black, but those from Europe are paler, suggesting coloration is variable depending upon environmental conditions. Details of adults and pupa drawn from Sasa (1978) and Yamamoto (1986,).
Male
AR 2.50-3.33. Wing length 2.9-3.2 mm, width 0.9 mm. LR 1.15-1.25, BR 2.2.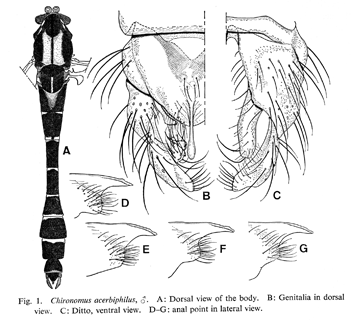
From Yamamoto 1986Head: Frontal tubercles, 22.5-35 µm long, 10-17.5 µm wide.
Ratio of palpal segments (µm) 59 : 64 : 203 : 199 : 254: P5/P4 1.28, P5/P3 1.25. 36-48 setae on clypeus.
Thoracic setae: Acrostichals: 8-10; dorsocentrals 13-23; prealars 6-11; supra alar 1; scutellar 22-36.
Legs: Note the measurements of Yamamoto (1986) are generally larger than those of Sasa (1978), and show the unusual feature that the antTa1 is shorter or only as long as AntFe, which is not seen in Sasa's measurements.
Proportions (µm) and ratios:
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
|
PI
| 1400
| 1220
| 1480
| 710
| 565
| 440
| 270
| 1.24-1.51
| 1.10-1.15
| 2.2
|
PII
| 1495
| 1250
| 675
| 390
| 310
| 210
| 185
| 0.51-0.58
| 1.10-1.25
|
|
PIII
| 1650
| 1495
| 990
| 560
| 445
| 265
| 205
| 0.65-0.70
| 1.07-1.13
|
|
Abdominal tergite IX with about 2-5 setae in individual pale spots. Hypopygium as in figure above. Anal point narrow and slender, slightly expanded at distal end.
Superior volsella figured by Sasa as Strenzke's D-type, and by Yamamoto as S-type. Strenzke (1959) described the German specimens (as C. crassimanus) as having a D-type Superior volsella, while Jablonska-Barna et al. (2012) record the Poland specimens as S-type; reaching just beyond tip of anal point. IVo apparently simple.
Female
Wing length 3.3-3.5 mm; width 1.1-1.2 mm; VR 0.81-0.87. LR 1.17-1.30.
Colour essentially as in male. Cercus black.
Head: Antennal segments (µm) 148 : 98 : 104 : 104 : 292; AR 0.62 (Sasa 1978)-0.64 (Yamamoto 1986). Frontal tubercle 15-38 µm long, 10-24 µm wide.
Ratio of palpal segments (micron) 66 : 66 : 202 : 208 : 260 (Yamamoto 1986) 40 : 50 : 200 : 200 : 250 (Sasa): P5/P41.25; P5/P3 1.25-1.29. 49-57 setae on clypeus.
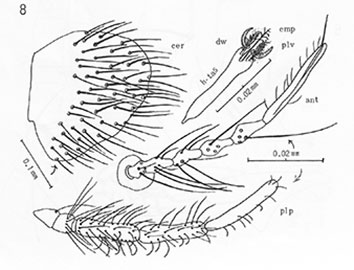
Cercus (upper left), Ta5 (upper right), Antenna (center), Palp (bottom)
of C. acerbiphilus (Sasa 1978)Thoracic setae: Acrostichals: 10-14; dorsocentrals 25-28; prealars 8-10; supra alar 1-2; scutellar 34-40.
Wing squama with 27-36 setae, bi- or tri-serial.
Leg proportions (micron) and ratios:
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| Ta5/Ti
|
PI
| 1600
| 1300
| 1630
| 715
| 600
| 465
| 285
| 1.17-1.30
| 1.20-1.25
| 0.21-0.23
|
PII
| 1645
| 1400
| 705
| 375
| 300
| 220
| 205
| 0.49-0.54
| 1.14-1.21
| 0.14-0.15
|
PIII
| 1775
| 1645
| 1035
| 570
| 480
| 290
| 225
| 0.61-0.67
| 1.04-1.12
| 0.13-0.14
|
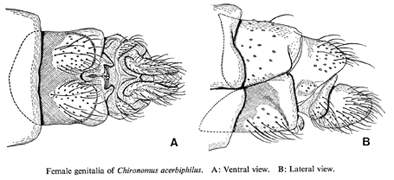
From Yamamoto 1986Genitalia: Apodeme of 8th sternum rounded caudolaterally, not joined mesally. Sternite of segment IX with 3-7 setae. Segment X wider ventrally and narrowing almost to a point dorsally, about 3.3 times longer than its widest point. Cercus somewhat "hat-like" with a gently rounded posterior margin, ventral margin longer than dorsal one.
Yamamoto (1986) shows a gently rounded base to ventral margin while Sasa (1978) shows a basal bulge.
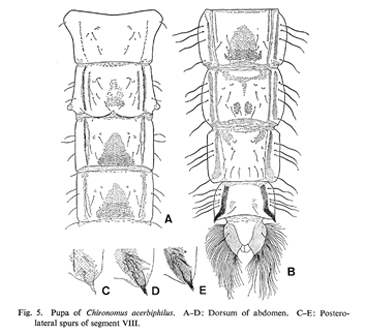
Pupa: (Based on Yamamoto 1986). Length 7.8-8.8 mm. Body dark brown. Cephalic tubercle acutely pointed with simple subapical seta. First and ninth terga practically without shagreen. Intersegmental membrane of V-VI segments and VI-VII segments with very weak centrally place shagreen.
The hook row is entire, but Jablonska-Barna et al. (2012) note that in Poland it is broken in some specimens; with 42-66 hooks in Europe.
Caudolateral spur of segment VII with 1-3 spines, most commonly with 2 (Sasa 1978).
From Yamamoto 1986Fourth instar larva: a small to medium sized plumosus-type larva, length 12.5-14.5 mm. Lateral tubules turn ventrally as described by Sasa (1978) for Japanese specimens. Ventral tubules well developed, anterior almost straight, posterior coiled.
Head capsule generally brownish; gula and frontoclypeus sometimes slightly to moderately darkened.
Labral setae simple. Salivarium long and narrow about 77 (71-83) µm long and 5.9 (5.5-6.2) times longer than wide.
Mentum width about half of ventral head length; c1 teeth relatively broad, with c2 teeth well separated and sharp (type III); Sasa (1978) shows a small notch near the top of C1, but this would only be seen where the teeth are not worn; lateral teeth sharp, with 4th laterals hardly reduced (type I) and 5th laterals slightly above the graduated level of the other lateral teeth.
Ventromental plates about 180-195 µm wide wide and 3.64-4.05 times wider than deep; separated by about 37-40% of mentum width; with about 47.6 (39-45) striae; VMR about 0.34 (0.21-0.45).
Pecten epipharyngis with about 22.5 (17-26) sharp graded teeth (although Yamamoto describes them as uneven).
Antenna with basal segment relatively long, about 3.5 times longer than wide; RO about 1/3 to almost 1/2 up from base of segment; AR about 2.35 (2.22-2.66); segment 3 short, may be same length as segment 5; relative length of segments (µm) 122.5 : 23 : 8 : 10 : 5.5.
Distance between antennal bases usually greater than that between the S4 setae, which are separated by about 75% of the frontoclypeus width at that point. S5 setae posterior to nearby RO.
Premandibles of type B1 with two narrow teeth, inner tooth about 3-3.5 times wider than outer tooth.
Mandiblewith 3rd inner tooth defined and darkened (type IIIC), about 14.3 (12-16() furrows on outer surface near the base; 13.2 (12-16) taenae in the Pecten mandibularis; Mdt-Mat 19-20 µm, MTR 0.31-0.33.
Cytology: 4 polytene chromosomes with the pseudothummi-cytocomplex combination AE, BF, CD, G.
Centromeres strongly heterochromatic and constricted. Pairing may occur betwen the centromeres of different chromosomes.
Arm G mostly paired, with BR near middle of arm and no nucleolus. Nucleolus developed in arm A.
A fixed asymmetrical pericentric inversion occurs on chromosome CD, transferring the proximal bands of arm D into arm C (Jablonska-Barna et al. 2010), or alternatively it may be a duplication of the CD centromere region (Wülker, pers. comm.), equivalent to those reported for the CD centromeres in some other pseudothummi-cytocomplex species such as C. dorsalis (=C. alpestris) (Kiknadze et al. 2008).
No polymorphism in studied Japanese, European or North American or populations.
aceA1: 1a-i, 7 - 9, 2d - 3, 12 - 10, 2c - 1k, 6 - 4, 13 - 19 with large nucleolus in segment 15.
aceB1: banding not clear, but probably 22-28 near centromere.
aceC1: 1 - 2, 10 - 3, 11 - 16, 22, 24 - 21, D(see below) (Jablonska-Barna et al. 2010).
aceD1: 1 - 3, 6 - 4, 7 - 9, 18f-a, 13 - 10, 17 - 14, 18g - 20 (Jablonska-Barna et al. 2010).
aceE1: 1 - 3e, 10b - 3f, 10c - 13 ie. as acidophilus, frommeri, whitseli, etc.
aceF1: 1, 12p - 11, 2 - 6 14 - 12p, 16 - 17, 10 - 7, 18 - 23 (Wülker, prelim).
aceF1: (alternate) 1-7, 17-16, 11-14a, 15-14b, 4-6, 9-8, 1-3, 10, 18-20 (clarified from Jablonska-Barna et al. 2010)
aceG1: BR near middle of arm.
Molecular data:
mtCO1: Sequence is in BOLD database and Genbank (DQ648201). Sequence of a North American specimen is also in the BOLD database. BOLD places it in a separate Bin (BOLD:AAL9507). This could simply be due to geographic isolation, but the differences in color and the fact that the Japanese specimens have a different nearest-neighbor Bin, suggest that they may be different species. A comparison of the base sequences shows that they differ at 35 bases in the Barcode region (below):
Base differences
USA A A T G G T T TA A T T C T A T T A C A C G G T A C T T G G A T G T A
Japan G G A A AC C C G GC C T C G C C G T T T A AA G T C A A A T C A C G