Chironomus novosibiricus Kiknadze et al. 1993In BOLD Bin: BOLD:AAW4007Originally described from Siberia, the presence of this species in China was demonstrated by Song et al. (2022).
This species has previously been placed in the camptochironomus-cytocomplex but molecular data from Guryev et al. (2001) points to the conclusion that this species has a different origin related to the thummi-cytocomplex (see below). Adult:
Based on Siberian specimens:Male:
Medium size, length 6-7 mm. AR-3.7. Thorax yellowish, bands and scutellum dark brown or black, postnotum dark; abdominal segments II-V with a broad blackish brown band at base and a distal yellowish band; segments VI-IX completely dark.
Legs yellowish, knees and tarsi slightly darkened, LR about 1.4.
Thoracic setae: dorsocentral - 29 (26-36); prealar - 6.5 (5-6); scutellar - 35 (33-49).
Unlike other species with the camptochironomus chromosome arm combination, this species has a normal hypopygium structure (below). 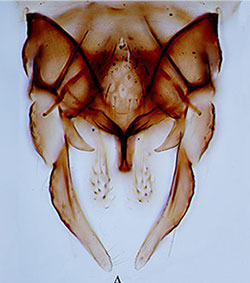
From Song et al. 2022Tergite IX with about 9-10 setae in several pale areas. Anal point broad, Superior volsella broad, E(d)-type of Strenzke (1959); Inferior volsella curved slightly outwards, longer than anal point and reaching about to middle of the gonostyle which is moderately broadened and narrows over distal third. Female and Pupa: - No information. Fourth instar larva: (From Russian specimens) Length about 11-13 mm. Some details can be determined from the figure (below). A bathophilus-type larva with short ventral tubules (not as long as the hind prolegs), anal tubules short without median constriction.
Gula darkened over most of its length, wider than mentum and lower at edges, widest at the posterior margin. Clypeus with dark central stripe. Antenna with RO in basal third, AR about 1.84; A1 about 3 times longer than wide.
Premandible with sharp teeth, inner tooth about 4 times wider than the outer, which narrows markedly along its length (Type B2). 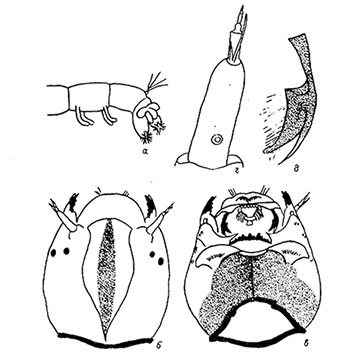
From Kiknadze et al. 1993Cytology: Three polytene chromosomes with a modified camptochironomus arm combination AB, CF, DEG. This is unlikely to have derived by the modification of the camptochironomus-cytocomplex. The cytology of the Chinese specimens is not known, but it is assumed that it is essentially similar to that of the Siberian populations. The following description is for those Siberian populations.
Chromosomes AB and CF are metacentric, while DEG is submetacentric with the centromere at the junction of the arms D and E. The single nucleolus is on arm G near the fusion with arm E; the arm also has two BRs near its distal end. The large puff on arm B is distal with dark bands (groups 7-10?) proximal. Siberian populations are polymorphic for inversions in all arms except F and G. novA1: 1a-2c, 12c-11a, 14h-13a, 4-9c, 2d-3, 10e-9d, 14i-19
novA2: 1a-2c, 6d-9c, 2d-3i, 12c-11, 9d-10, 6c-4a, 13a-19
novB1: not mapped
novB2: inversion of about half the arm with one break about 12 bands from the centromere.
novC1: 1a-3c, 15e-14, 10-8, 12-13,11a-h, 6b-4, 6gh, 17a-16, 7d-a, 6f-c, 17b-22g
novC2: 1a-3c, 4-6b, 11f-a, 13-12, 8-10, 14-15a, 6gh, 17a-16, 7d-a, 6f-c, 17b-22
novD1: 1a-3, 18d-a, 8-10c, 14hg, 19e-18e, 17-15, 14f-10d, 4-7, 19f-24
novD2: 1a-3, 18d-a 8a-10c 14h-10d 4-7, 15-17, 18e-24
novD3: not mapped
novE1: 1a-c, 3f-4, 1d-2, 10b-5, 3e-a, 10c-13
novE2: 1a-c, 10b-5, 3e-a, 2e-1d, 4-3f, 10c-13
novF1: 1a-d, 18-19, 7-8d, 14-11d, 17-15, 10d-a, 8e-9, 6e-1e, 20-23
novG1: not mapped
(from Kiknadze et al. 2016) 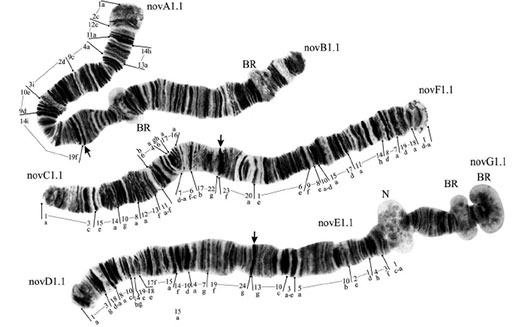
Polytene chromosomes of European C. novosibiricus.
From Kiknadze et al. (2016)Found: Type locality - Siberia, Novosibirsk, Novosibirsk Provence
China - Zhenxing, Dandong, Liaoning (40.1217°N, 124.392°E).
Russia – Numerous localities in the Urals; Novosibirsk Provence; Tomsk Provence; Altai Territory and Republic of Altai; Republic of Tyva; Kazakhstan.> DNA sequence:
MtCOI: As noted above there is a sequence in the BOLD database.
Additional sequence for mtCOI from Siberian populations is in GenBank and also in the BOLD database, including from Guryev et al. (2001), who also studied sequence for gb2B. This latter sequence casts doubt on any relationship between C. novosibiricus and the species of the camptochironomus-cytocomplex as the latter lack the intron of the gb2B gene, while C. novosibiricus contains the type-2 intron found in many thummi-cytocomplex species. On the basis of this Guryev et al. (2001) suggested that C. novosibiricus is instead derived from a species with a modified thummi-cytocomplex arm combination - probably C. tuvanicus, to which it is closely related, or a common ancestor to both. [ Return to Index | Go to References ] |