6b. Chironomus 'paracastaneum' n. sp.This species is recognized as close to C. 'castaneum' on the basis of the cytology and mtCOI sequences, and also close to C. sp.NZ9. Along with C. 'castaneum' and C. sp. NZ9. (see under C. sp.NZ9 for further detail) Adult: Two males, one a somewhat incomplete reared male were available for study. The name C. 'paracastaneum' is suggested for this species and the adult male from Lake Hawea, Otago, South Island, NZ.48.1 male 1 (7.I.1974) has been chosen as the potential holotype since there is also DNA barcode data for this specimen. Colouration not recorded before DNA extraction and slide mounting. Frontal tubercles present, length about 40-69 µm long and 16-18 µm wide.Palpal proportions (micron): 78 : 56 : 280, 278 : 420. About 26 clypeal setae. Thoracic setae: at least 10 acrostichals; 22 dorsolaterals; 6-7 prealars; 1 supra-alar; 27 scutellar in two rows (10 in anterior row, 17 in posterior row). Leg lengths (micron) and proportions:
Abdomen with somewhat triangular (Don Forsyth referred to them as 'bell shaped') brown markings on the tergites, narrowing towards the posterior margin on segments II-IV, then covering most of the tergites. TIX with 9 setae in a single pale area. 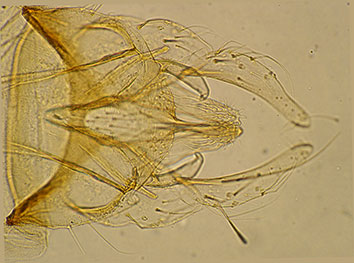 Hypopygium of the proposed type male. Anal point narrow at base and widening a little at the distal end. Superior volsella closest to the E(h) type of Strenzke (1959); Inferior volsella reaching about to the end of the anal point or a quarter to a third along the gonostylus, with simple setae. Gonostyle moderately swollen and narrows markedly from about the distal third to half; about 6+1, 7+1 setae at tip. Additional data from the other male: AR about 3.7. Wings damaged LR about 1.33-1.52, anterior tarsi without a beard (BR about 1.7). Female: There is data for a female collected at the same time and from the same site as the proposed type male. It is included here although there is no proof that it belongs to this species (C. forsythi was also present at the location, but the leg ratios are more similar to those of the type male than to those of C. forsythi females). Wing length 5.37 mm; wing width: 1.14 mm; VR 1.09.AR: 0.35; flagella length (micron): 225 : 130 : 155 : 140 : 230, A5/A1 1.02. Palpal segments (micron): - : - : 105 : 275 : 301; P5/P4 1.35; P5/P3 2.86. Clypeal width 1.8 times antennal pedicel; 37 setae. Thoracic setae: About 16 acrostichal; 4-5 humeral; 25-29 dorsolateral (30-33 dl+humls); 8 prealars; scutellars in two approximate rows - anterior row about 15, posterior row about 19 (34 total) setae. Leg lengths (micron) and proportions:
Colour of abdominal tergites undetermined but apparently over most of the segment. GcIX with 3 setae; segment X appressed to cercus, crescent shaped, about 280 µm long and 5.9 times longer than its greatest width, with 16 setae. Cercus (below) with rounded outline; ventral margin longer. 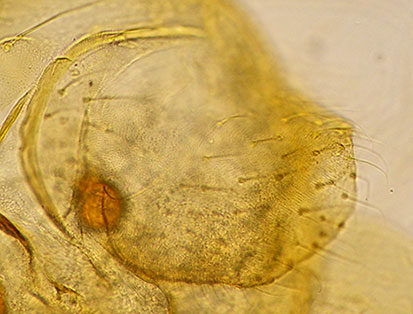 Cercus with closely applied segment X (at left). Pupa: Length about 9.4 mm. Pedes spurii B developed on segment II, and about 55 hooks on posterior margin. Pedes spurii A well developed on segment IV. Fourth instar larva:: A salinarius-type larva, length 12.7-17.2 mm (female); 12.3-14.8 mm (male). A larva from Winton showed a small bump for the posterior VT (about 0.06mm). Anal tubules short (200-400 µm) and rounded, only about 2-3x (1.8-3.2) longer than wide. Cytology: 4 polytene chromosomes, with the pseudothummi group arm combination (BF, CD, EA, G). camA2: 1a-e, 2e - 3, 1f - 2c, 10 - 11, 2d, 8 - 9, 3f-i, 12c-a, 4 - 7, 13-19 i.e. as casA2 camA3: 1a-e, 12a-c, 3i-f, 9 - 8, 2d, 11 - 10, 2c - 1f, 3 - 2e, 4 - 7, 13 - 19 from camA1 (Winton, & also Poutu as casA3) camB1: Nucleolus just distal to 4 characteristic bands (grps 24-26), and puff with distal dark bands (grps 8-7) near distal end. i.e. as casB1 camB2: Inversion of distal third of arm, as nzlB3, bringing puff and proximal dark bands (groups 7-8) close to nucleolus. i.e. as casB2 camB3: Inversion of region of the nucleolus, from camB2 (different breakpoints to the difference of camB1 to forB2) camC1: groups 4-3 about 1/3 from distal end, with groups 6-5 distal to it. i.e. as casC1 camC2: simple inversion of distal third from camC1. (in NZ.67.1 Em#1) camD1: as casD1 camD2: as casD2 camE1: 1 - 3e, 10b - 3f, 10c - 13 i.e. as oppE1, etc., casE1 camE2: 1a-e, 3f - 10b, 3e - 2a, 10c - 13 i.e. as casE2 camF1: 1 - 2a, 10 - 2c, 15c - 11a, 2b, 15d - 23 i.e. as casF1, oppF3, forF1 camF2: (approx.) 1 - 2a, 10 - 9c, 4 - 9b, 3 - 2c, 15c - 11, 2b, 15d - 23 as forF2 camG1: with small subterminal nucleolus camG2: from camG1 by a smaller inversion around the median BR than that differentiating camG1 from forG1. (in NZ.67.1 Em#1) Click here to see chromosome BF heterozygous for the inversion in each arm. Known Localities The evidence for this species comes from the cytology and the MtCOI sequence. The cytology and the MtCOI sequence suggests that there may also be hybridization between C. 'paracastaneum' and C. analis, since at least one specimen has been found with the C. 'campanello' cytology, but the C. analis CO1 sequence.
|
Modified: 22 September 2023
Access: Unrestricted
Copyright © 2000-2023, Jon Martin.