Chironomus vitellinus Freeman 1961Placed as a synonym of C. javanus by Chaudhuri et al. 1992, but this seems to be incorrect as the anal point of the adult males is expanded at the end; there is a difference in tooth number of the larval premandible and there are significant differences in the BARCODE sequences. In BOLD Bin: BOLD:6924. Adult Further description: Wing length 2.81 (2.54-3.08) mm, width about 0.61 mm, VR about 1.05; 18 (11-26) setae in squamal fringe; 2 SCf on brachiolum. Head: AR 3.04 (2.78-3.46). Frontal tubules about 40 (30-45.5) µm; 16.5 (11-21) clypeal setae Palpal proportions (µm) 44 : 45.5 : 157 : 196 : 275; P5/P4 1.38-1.45; P5/P3 1.78-1.82 Thoracic setae: 3 (0-5)acrostichal; 8.4 (5-15) dorsocentral; 4.1 (3-7) prealar ; supraalar: 0.67 (0-2) in anterior row, 10.32 (5-16) in posterior row, total 10.7 (10-16) scutellars. Leg lengths (micron) and proportions:
Tergite IX with 8.5 (7-10) setae in individual patches. Anal point narrow but expanded at distal end, narrow in lateral view (see figure below); Superior Volsella closest to E(g)-type of Strenzke (1959); Inferior Volsella short and very stout, not reaching the end of the anal point, but to about 1/3 of the gonostylus length, with simple setae; gonstylus quite swollen (not always as much as in the Tokunaga figure below) and decreasing markedly over posterior third. 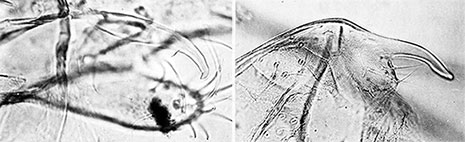 Superior and inferior volsellae (left) and lateral view of anal point (right) of paratype male of C. vitellinus
Wing length 2.81 (2.08-2.93) mm, width 0.83 (0.66-0.96) mm; VR 1.09 (1.07-1.11); 2 Scf on brachiolum; 15.6 (13-16) setae in squamal fringe. Head: Frontal tubercles present 28.1 (7.5-35) micron long and 1.0-2.8 times longer than wide. Antennal segments (micron) with percentage neck in brackets: 169 (28) : 115 (46) : 124 (49) : 126 (49) : 200; AR 0.37 (0.33-0.43); A5/A1 1.10 (1.07-1.17). Palpal segments (micron): 54 : 50 : 180 : 220 : 335; P5/P4 1.53; P5/P3 1.86. Clypeus heart-shaped, about 1.28-1.55 wider than antennal pedicel; abt 23 (16-39) setae. Thoracic setae: 11.5 (9-16) acrostichals; 3.6 (3-5) humerals, mostly linear but may be grouped (e.g. as a triangle); dorsocentrals - 15.2 (9-260); 18.7 (13-30) including the humerals (lower in Pacific Islands); 5 (4-8) prealars; 2.2 (0-6) anterior row and 10 (8-13) in posterior row (total 8-19) for scutellars. Leg lengths (microns) and proportions as follows:
GcIX with 3.7 (2-6) setae; segment X usually a half-oval 91-177 µm wide and 2.99 (2.1-5.36) times longer than greatest width, with about 11.4 (10-13) setae. Sasa & Hasagawa (1983) note that the cercus is roughly rhombic, 112x152 µm; usually with a ventral basal bulge. Pupa, Fourth instar larva and cytology: see immatures, etc. The adult male was described from Thailand by Hashimoto et al. (1981), and all stages for specimens from Japan (Hasagawa and Sasa 1987) and from Micronesia by Tokunaga (1964), all as C. javanus. The expansion at the tip of the male anal point, the setae on TIX and the 7 toothed premandible clearly show that these descriptions refer to the present species (and supported by the BARCODE data which show that C. vitellinus is more common and more widely distributed). Molecular: Mitochondrial CO1 barcode sequence appears to be in the BOLD database under the name C. javanus and is much more numerous than those of C. javanus.  Found: AUSTRALIA Type locality - Darwin, Northern Territory, Australia; |
.jpg)
Modified: 1 June 2026
Access: Unrestricted
Copyright © 2010-2026, Jon Martin.