C. tepperi immaturesEgg mass: The egg mass is a typical thin sausage shape. Factors affecting the size of egg masses, and the number of egg masses laid by fed or unfef females was given in Martin & Porter (1977). Fed females laid more egg masses (up to 5 or 6 of decreasing size, and often reducing fertility), while unfed females rarely laid more than one egg mass. It has also been found that the number of eggs in an egg mass is correlated with the wing length of the female (correlation coefficient r = 0.922). Fourth instar larva a medium sized (9.8–14.8 mm)halophilus/bathophilus, variable; Ventral tubules sometimes moderately developed (bathophilus) (ant. 0.33–0.83 mm; post 0.67–1.17 mm) i.e. posterior pair longer, but sometimes only posterior pair present (halophilus) (see below). Gular region pale, frontoclypeus usually not darkened, but occasional very slight darkening. Hind prolegs reduced quite markedly near the tips (see below).
Mentum (Fig. c) with 4th lateral reduced slightly, sometimes to about the level of the 5th lateral (type I-II), and c2 teeth notches on the c1 tooth (type IIA, sometimes tending towards type III).
Ventromental plates (Fig. d) well separated (76-96 µm) by about 44-46% of mentum width, with about 26-32 striae; VMR about 0.27. Pecten epipharyngis (Fig. a)with about 12-17 teeth.
Premandible (Fig. b) with inner tooth about 2-5x wider than outer tooth, coming to a fine point (type B1).
Basal segment of antenna (Fig. c) relatively long and narrow, about 3.7-5 times longer than wide, ring organ about one third from base of A1; AR about 2.59 (2.43 - 2.74; A2/A1 about 0.21 (0.20-0.21); antennal proportions 137 : 28 : 8 : 10 : 7 .
Mandible (Fig. f) with third inner tooth completely formed (type IIIC), with about 12-16 furrows on outer surface near the base; MTR about 0.35. 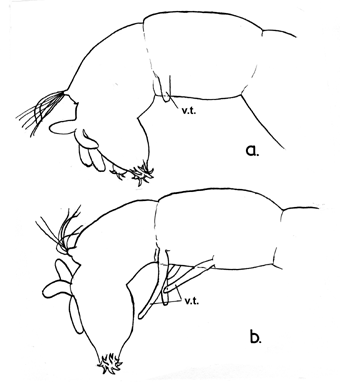
Terminal abdominal segments of C. tepperi
showing different degrees of development of the ventral tubules seen in various populations
Note also the relatively abrupt narrowing of the prolegs (best depicted in 'a').
From Martin, J., M.Sc. Thesis, The University of Melbourne, 1961.Cytology: 4 polytene chromosomes with the pseudothummi arm combination AE, BF, CD, G. All chromosomes closely paired.
Arm G polymorphic for a subterminal nucleolus, with 2 Balbiani rings. Nucleolus in arm F.
No inversion polymorphism known. Irradiation experiments have revealed that the male sex determiner (MD) is located in the proximal region of arm A, between 3f and 19e, with 8% recombination between the MD and the centromere (Martin, 1980).
tepA1: 1a-e, 11 - 10, 2c - 1f, 3e - 2d, 8 - 9, 3f-i, 12c-a, 4 - 7, 13 - 19 as oppA4
tepB1: Bulb and distal dark bands about 1/4 from end of arm. as oppB2
tepC1: Characteristic band groups 3-4, with 5 distal, near distal end as 'pseudoppositus'C1
tepD1: 1-2f, 9b-e, 3d-a, 10d-12, 18g-a, 2g-i, 16-14h, 19c-a, 8-3e, 10a-c, 17a-f, 9a, 13-14g, 19d-24
tepE1: 1 - 3e, 10b - 3f, 10c - 13 as oppE1, etc.
tepF1: 1 - 2a, 10 - 2b, 11 - 23 as oppF1 Pupal exuvia pale, length about 8.9 mm (6.6-9.3 in females, 6.7-10.6 in males). Cephalic tubercles about 65-70 µm long, width at base 40-58 µm, subapical seta about 50 µm.
Large pedes spurii B on segment II, that on segment III a relatively large patch of spines. Large pedes spurii A on segment IV, with smaller patches of spines on segments V and VI.
About 60-77 recurved hooks on segment 2; about 78-120 taeniae along the edge of the swim fin, mean number higher in females (abt 103 cf. 93 in males).
Caudolateral spur of segment VIII with about 6 spines (3-8), closely applied. The salivary gland chromosomes were described by Martin (1974); the nucleoli and location of C-bands by Lentzios & Stocker (1979) and Lentzios et al. (1980). Kalisch (1981); Kalisch & Hägele (1981, 1982) and Schmitz (1982) produced photographic maps of the polytene chromosomes based on surface-spread chromosomes. See also C. tepperi adult; C. oppositus larva.[ Return to Index | Go to References ] |