 |
AUSTRALIAN CHIRONOMUS
SPECIES |
Compiled by Jon Martin
Balwyn North, Victoria, 3125, Australia
When Freeman (1961) revised the Australian Chironomidae he recognized six species of the genus Chironomus (s.s.): C. alternans Kieffer, C. australis Skuse, C. nepeanensis Skuse, C. tepperi Skuse and C. vitellinus Skuse. However, even at that stage it was known that adults of C. alternans could be produced from distinct larval types, and with quite distinct polytene chromosomes, which appeared to be distinct species. Bugledich et al. (1999) 'Family Chironomidae' from Zoological Catalogue of Australia, recognize 12 species of Chironomus in Australia, with another nine species that are Incertae cedis. Only two of these latter nine, which are currently nomena nuda, are considered here. Here 25 species are considered although 11 are only manuscript names and not formally described. There are a further 3 species from Norfolk Island.
There is also one species of Einfeldia in Australia, E. australiensis (Freeman) which Freeman placed in Chironomus (Xenochironomus). This species is included here as the relationship of Einfeldia to Chironomus has been subject to much discussion. | This listing is also available in pdf format (7.5 MB)(Updated 11 April 2024).One aim in these studies has been to evaluate as many characters of adults, pupae and larvae as possible in order to evaluate which ones might prove useful for separating the species that can only be recognized at present by the banding patterns of the larval polytene chromosomes or the BARCODE sequence. In general, the morphological terminology used in this document follows Sæther (1980), Webb & Scholl (1985) and Vallenduuk & Moller Pillot (1997).
Most of these studies were carried out in the Zoology and Genetics Departments of The University of Melbourne.
1. Adult morphologySpecies with an Australia-wide distribution
Chironomus cloacalis Martin, 1971
Chironomus 'februarius' - currently nomen nudum, but described in Martin 1966
Chironomus 'pseudoppositus' - currently nomen nudum, but described in Martin 1966
Chironomus tepperi Skuse, 1889 Species with a tropical or Pacific distribution
Chironomus bicoloris Tokunaga, 1964
Chironomus circumdatus (Kieffer, 1916)
Chironomus nepeanensis Skuse, 1889
Chironomus 'orientalis'
Chironomus pallidinubeculosus Tokunaga, 1964
Chironomus magnivalva Kieffer, 1917
Chironomus vitellinus Freeman, 1961 Species with a north eastern distribution
Chironomus 'queenslandicus' - manuscript name
Chironomus sp. (bathophilus type)
Chironomus n.sp. Qld
Chironomus(?) blandellus Kieffer, 1906 Species with a south eastern distribution
Chironomus australis McQuart, 1847
Chironomus duplex Skuse, 1889
Chironomus 'jacksoni' - manuscript name
Chironomus maddeni Martin & Cranston
Chironomus oppositus Skuse, 1889:
form connori
form oppositus
form whitei
Chironomus 'timmsi' - manuscript name
Chironomus 'tyleri' - manuscript name (formerly C. oppositus f. tyleri)
Chironomus 'tasmaniensis' - manuscript name
Einfeldia australiensis Species with a western distribution
Chironomus nepeanensis Skuse, 1889
Chironomus occidentalis Skuse, 1889
C. oppositus f. edwardi Species from Norfolk Island
Chironomus 'februarius' (?)
Chironomus species NI 1 (may be as Chironomus samoensis)
Chironomus species NI 2 The description of Chironomus alternans Walker and the synonyms listed by Freeman (1961) are also included, although they are considered as uncertain. Further description is provided for those where the types are still available:
C. alternans
C. januarius
C. subdolus
In the descriptions of adult males reference is made to the types of superior volsella shape as recognized by Strenzke (1959). This is a helpful initial classification, but experience has shown that the types are not discrete, but are part of a continuum. The three categories as described by Strenzke are:
S-type: The SVo is shoe shaped, i.e. it is drawn out distal-medially into a broad, rounded lobe (Fig. a-c, below) (Strenzke's figure suggests the most distal point will be at the toe of the shoe).
D-type: The SVo is ribbon-like: distally it may have a weakly thickened shoulder (Fig. d, below) (most distal point is not at the internal margin), or bent in a shallow sickle-shape (Fig. e-f, below).
E-type: The SVo has the form of an elephant's tusk; distally it is sharply graded to a point, or with an expanded knob (Fig. g-i, below) (line from base to most distal point goes outside the limits of the SV).

Key to Adults
Adult males:
| 1a | Yellow/green species. Anal point stout | 2a | | 1b | Anal point longer or tubular | 3a | | | | | | 2a(1a) | Anal point bulky and strongly turned down, abdomen greenish | Einfeldia australiensis | | 2b | Anal point not bulky or strongly turned down, abdomen dark | C. 'queenslandicus' | | | | | | 3a(1b) | Wing with dark spot over cross vein and with fuscous clouds, particularly in cell R5 | C. pallidinubeculosus | | 3b | Wing with at most darkening of cross vein | 4a | | | | | | 4a(3b) | Cross vein strongly darkened, LR about 1.5, wing length 4-5 mm; ant TA4 longer than Ta3 | C. nepeanensis(northern),C. atchleyi(southern) | | 4b | Cross vein not strongly darkened | 5a | | | | | | 5a(4b) | Legs shorter than usual, posterior LR about 0.5; male hypopygium enlarged, anal point deep and laterally flanged | 6a | | 5b | Posterior LR about 0.7 or higher; male hypopygium of normal type | 7a | | | | | | 6a(5a) | Thorax dull grey, pits of dorsocentral setae very distinct; inferior volsella of male very swollen | C. tepperi | | 6b | Thorax greenish with brown stripes, pits normal; inferior volsella of male not swollen | C. magnivalva | | | | | | 7a(5b) | Wing length greater than 5 mm; anterior tarsi of males bearded | 8a | | 7b | Wing length less than 5 mm; anterior tarsi of males bearded or unbearded | 10a | | | | | | 8a(7a) | Distributed on east of the continent, including Tasmania | 9a | | 8b | Distributed on the west of the continent, including Northern Territory | C. occidentalis | | | | | | 9a(8a) | Mid femur more than 1.12 x the anterior femur (likely to be) | C. duplex | | 9b | Mid femur shorter than the anterior femur (likely to be) | C. australis | | | | | | 10a(7b) | Blackish species. Male with bearded anterior tarsi and spatulate superior volsella | C. 'timmsi' | | 10b | Paler species, at most brown. Male tarsi not bearded | 11a | | | | | | 11a(10b) | Thorax pale, anterior femur and tibia pale, tarsi with dark pattern | 12a | | 11b | Thorax pale to brown, legs unbanded (mainly members of the 'C. alternans' group) | 13a | | | | | | 12a(11a) | Tarsi darkened at both apex and base, anal point narrow in lateral view | C. vitellinus | | 12b | Tarsi darkened only at distal ends, anal point larger and narrowed at base, LR above 1.80, anterior tarsus 5 at least 0.35 times the length of the tibia, SV often beaked | C. 'orientalis' | | | | | | 13a(11b) | Wing length over 4 mm. | 14a | | 13b | Wing length less than 4 mm | 15a | | | | | 14a(13a) | Abdomen generally greenish with saddle markings on the anterior segments, gradually extending in length from about 5th tergite to occupy most of segment by 8th tergite (AR 2.9-3.2) . LR 1.53-1.66 | C. 'tyleri' | | 14b | Abdominal tergites generally dark with pale posterior band, but may have saddle spots;.(AR 3.3-4.25); wing length up to 4.6 mm. LR 1.44-1.78 | C. cloacalis in part | | 15a(13b) | | C. oppositus type | | 15b | Abdominal tergites generally dark, may have pale posterior band, saddle spots, or dark band across the centre of the segment | 16a | | | | | 16a(15b) | as 14a, but wing length from 2.6 mm | C. cloacalis in part | (Abdominal tergites may be as C. cloacalis, or may have dark band across the centre of the segment; AR 2.9-3.4; LR 1.56 - 1.65.), C. 'pseudoppositus' (Abdominal tergites as in C. cloacalis AR 2.5-3.1; LR 1.5-1.8; superior volsella less curved than in other members of the oppositus-group)
Abdomenal tergites with saddle spots: C. oppositus (LR 1.5-1.8; wing length 2.8-3.6, C. maddeni (AR 2.55-2.86; LR 1.6-1.8; wing length 2.4-3.36 mm), C. 'jacksoni' (AR 2.4-3; LR ); Chironomus 'tyleri' (AR 2.9-3.2; LR 1.53-1.66; wing length 4.3-4.5 mm). Norfolk Island species
| 1a | LR about 1.5, superior volsella closest to Strenzke's type D(d); may be near C. samoensis. LR is lower than C. nr. samoensis from other regions, but in range of C. samoensis itself). | Species 1 | | 1b | LR about 1.27, superior volsella closest to Strenke's type D(f), but appears to have a flattened end | Species 2 | | Adult females
No key to the females currently exists but some of the characters common to both sexes can apply, as well as additional characters from the antennae, the shape of segment X and the shape of the cercus. Also, the dorsocentral setae begin more anteriorly than in males, the additional setae anterior to the parapsidal suture are the humeral setae and their arrangement can be useful as a taxonomic character.
2. Larval types and KaryotypesIn the following descriptions, reference is made to the mentum and mandible types originally devised by Webb & Scholl (1985), Vallenduuk & Moller Pillot (1997), and Proulx et al. (2013). These classifications were made for relatively small numbers of species, but with the much larger number of species, such as in the North American fauna, they do not cover all the variability seen in these characters and so further modification has been necessary. The MENTUM TYPE is defined only by the degree of development of the 4th lateral teeth:

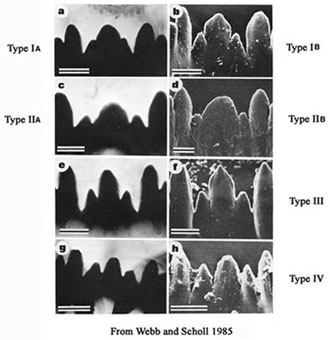
Type I - height in same line as the rest of the lateral teeth;
Type II - 4th laterals reduced, height about equal to that of the 5th laterals;
Type III - 4th laterals further reduced, height less than that of the 5th laterals.
The mentum may be further classified by the characters of the CENTRAL TRIFID TOOTH:
Type IA - c2 teeth only partially separate from c1, i.e. as shoulders on c1 (figure a).
Type IB - c2 teeth slightly more separated (figure b).
Type IIA - c1 broad, c2 teeth distinctly separated (figure c).
Type IIB - c1 very broad, c2 less separated (figure d).
Type III - c1 tooth relatively narrow and much higher than the separated c2 teeth (figs e and f).
Type IV - c2 teeth well separated, not much lower than the relatively narrow c1 tooth (figs g and h).
The MANDIBLE TYPE is defined by the degree of darkening and separation of the 3rd inner tooth:
It seems better to consider the two characters separately.
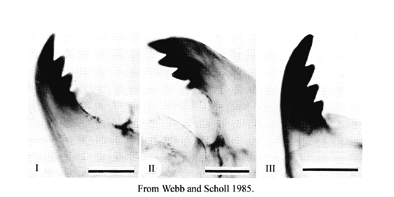 This figure shows IA; IIB; IIIC respectively
Separation:
Type I - tooth fused
Type II - tooth partially free
Type III - tooth completely separated
Color:
Type A - tooth pale
Type B - some degree of pigmentation
Type C - as dark as other inner teethMandible length and Mdt-Mat:
Hirvenoja and Michailova (1998) illustrated that the distance between the tip of the dorsal tooth and the apical tooth could differ between related species (Mdt-Mat) (blue line in figure below).
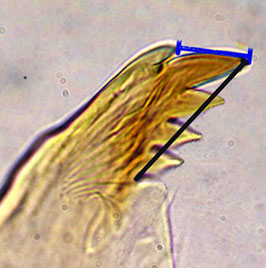
However for different sized species it may be preferable to divide this value by the length of the inner teeth (black line in figure above) to obtain the MTR.Ventromentum
There are several measures that can be made from the ventromental plates including distance between inner margins of the plates (VPA) and the number of striae. Two others need some explanation: VENTROMENTAL PLATE RATIO (VMR) - ratio of the width of the marginal region of ventromentum (usually seen as a granular band under light microscopy) (a in figure below) to the distance from the anterior margin to the base of the striae (b in figure below).
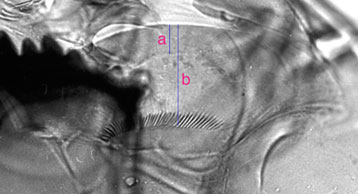
VMR = a/b
'b' also serves as a measure of the depth of the VM for the ratio of length to depth of the VM plates.
VM length (VML) is measured directly from inner margin to outer margin:
This seems preferable to the sometimes used 'horizontal length' which can be subject to parallax error and to the effects of rotation of the plates under pressure during slide mounting.PECTEN EPIPHARYNGIS -
Proulx et al. (2013) recognised 4 types of PE in North American species. These are useful if the teeth are not worn down, as they often are in older larvae. Type D does not occur in any known Australian species.
Type A - fine sharp rather uniform teeth.
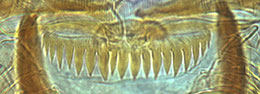
Type B - teeth broader but still sharp. Sometimes with one or two fine smaller teeth interspersed.
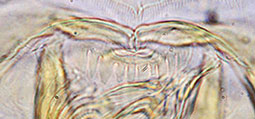
Type C - rounded and rather uniform. Worn type B teeth may be mistaken for this type.
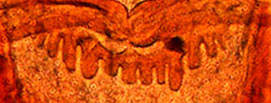
Type D - rounded teeth with smaller teeth interspersed (generally found in the subgenera Lobochironomus or Chaetolabis).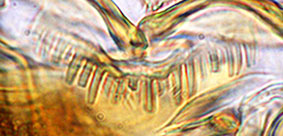 PREMANDIBLE -
Assessment of the premandible can be difficult due to quite excessive wear and distortion in mounting. Where they are in good condition and lying flat, they can be categorised into 5 groups based on the relative widths of the teeth and whether they come to a sharp or a broad point. The teeth are measured as close as possible to their bases.
Premandible types
Type A: Both teeth narrow and coming to a fine point.
Type B: inner tooth moderately broad, about 3-5 times wider than, outer tooth. This can be split into 2 subgroups:
B1 � both teeth come to relatively fine points.
B2 � the inner tooth comes to a relatively broad point.
Type C: Both teeth are moderately broad, coming to relatively broad points.
Type D: Inner tooth very broad, outer tooth moderately broad.
Type E: Both teeth very broad and often quite short.
There is also at least one species (C. sp. Florida) recorded with a five toothed premandible similar to that of C. javanus.
Species of Kiefferulus also have a five-toothed premandible, but it is broader. ANTENNAL RATIO (AR) - length of A1 divided by the sum of legths of A2-A5, measured on the basis of the chitinised part of each segment only, as the inter-segmental regions can extend to different extents during mounting. For this reason the AR will usually be higher than that obtained by simply measuring the total length from base of A2 to tip of A5. Relationship on the Frontoclypeus (FC) of the distance between antennal bases and distance between S4 setae
This character gives some indication of the shape of the anterior region of the FC: the amount and extent of the narrowing at the anterior end near the antennal bases, and where the S4 setae are in relation to the broadening of the clypeus (see figure below). This relationship can be further characterized by the distance of the S4 setae from the margins of the FC � most easily expressed by the fraction of the FC width between the two S4 setae. This has two components: how far the setae are from the FC margin, and how close they are to the widest point of the FC.

Frontoclypeus with approximately equal distance between antennal bases and S4 setae.
Note also the barely visible 'ring organ' of Yamamoto et al. (2015), (more obvious at top) immediately opposite the S5 setae. This is a characteristic of species of Chironomus.Larval types referred to in Key and in descriptions are defined here. Provisional Key to 4th Instar Larvae
| 1a | Larger larvae, length up to about 23 mm. (Either thick bathophilus-type larvae with large, very darkly pigmented heads, or a slim plumosus-type larva with a small, pale head capsule) | 2a | | 1b | Medium sized larvae, length up to about 19 mm, head length less than 400 mm. | 5a | | | | | | 2a(1a) | Head capsule heavily darkened, bathophilus-type | 3a | | 2b | Head capsule pale, except NT and Vic where slight darkening of the gula & frontoclypeus occurs, slender plumosus-type | C. nepeanensis (northern/western); C. atchleyi (southern). | | | | | | 3a(2a) | Distributed on east side of the continent only (NSW, VIC, SA, TAS) | 4a | | 3b | Distributed on west of continent only (WA & perhaps NT) | C. occidentalis | | | | | | 4a(3a) | Teeth of mentum rounded (when not worn), 4th inner tooth of mandible type B. Four polytene chromosomes | C. australis | | 4b | Teeth of mentum square sided and pointed, 4th inner tooth mandible type C. Three polytene chromosomes, inversion polymorphism | C. duplex | | | | | | 5a(1b) | Larvae with lateral tubules (species often polymorphic) | 6a | | 5b | Larvae without lateral tubules (species often polymorphic) | 12a | | | | | | 6a(5a) | Larvae of the true plumosus type (posterior ventral tubules coiled) | 7a | | 6b | Larvae of the melanotus- or semireductus- type (posterior ventral tubules may be bent) | 16a | | | | | | 7a(6a) | Head capsule generally quite heavily darkened on both frontoclypeus & gula | 8a | | 7b | Head capsule darkened on gular region only | 10a | | | | | | 8a(7a) | Both gula and frontoclypeus quite heavily darkened | C. cloacalis (majority) | | 8b | Gula and frontoclypeus slightly to moderately darkened, tropical species | 9a | | | | | | 9a(8b) | c2 teeth of mentum sharp and well separated (type III), 3rd inner tooth of mandible type IA/B | C. 'orientalis'. | | 9b | c2 teeth of mentum only partially separated (type IB), 3rd inner tooth of mandible type IIIB | C. magnivalva | | | | | | 10a(7b) | Lateral tubules tending to be more ventrally placed, premandible with 7 teeth | C. vitellinus | | 10b | Larvae not as above, premandible with the normal 2 teeth | 11a | | | | | | 11a(10b) | Head capsule often completely pale, but sometimes slightly darkened at posterior of gula, basal segment of antenna less than 4 times as long as wide | C. pallidinubeculosus | | 11b | Basal segment of antenna more than 4 times as long as wide: | 12a | | | | | | 12a(11b) | Larvae of the halophilus-type (i.e. VT reduced, often only the posterior pair ), hind prolegs sharply narrowing | C. tepperi (part) | | 12b | Larvae with VT of long or moderate length | 13a | | | | | | 13a(12b) | Larvae of the plumosus-type | 14a | | 13b | Larvae of the bathophilus-type | 18a | | | | | | 14a(13a) | Gula darkened at least on posterior half | 15a | | 14b | Gula pale or only slightly darkened on posterior third | 17a | | | | | | 15a(14a) | Gula dark, centre tooth of mentum of type III | C. circumdatus | | 15b | Gula moderately darkened, centre tooth of type IB | pale headed form of C. cloacalis | | | | | | 16a(6b) | Lateral tubule over 200 micron; inner margins of ventromental plates separated by only about 0.2 of mentum width; AR abt. 1.40-1.64; PE of type D | C. 'queenslandicus' | | 16b | Lateral tubules often less than 200 micron; ventromental plates separated by about 0.3 of mentum width, AR >1.9; PE usually type B | some C. oppositus f. oppositus, C. 'edwardi' and C. 'tyleri'. | | | | | | 17a(14b) | Mentum of type II; Central trifid tooth type III | C. 'februarius' | | 17b | Mentum of type I; Central trifid tooth type III | C. bicoloris | | | | | | 18a(13b) | Basal segment of antenna 3 to 3.5 times longer than wide; four polytene chromosomes | 19a | | 18b | Basal segment of antenna about 4 times longer than wide; three or four polytene chromosomes
(Chironomus n. sp. Qld might fit in this couplet, but there is no information on larval type or chromosome number) | 20a | | | | | | 19a(18a) | Gula and frontoclypeus pale; antennal segments A2/A1 less than 0.3 | C. 'timmsi' | | 19b | Gula and frontoclypeus at least slightly darkened, A2/A1 more than 0.3
Accurate identification only possible from examination of the polytene chromosomes. | most members of C. oppositus complex, C. 'pseudoppositus', C. maddeni and C. 'jacksoni' | | | | | | 20a(18b) | Basal segment of antenna about 4 times as long as wide; three polytene chromosomes | C. sp. Bakers Beach | | 20b | A4 hardly longer than A3, four polytene chromosomes
(some specimens of C. tepperi may key to here if they have relatively well developed ventral tubules. They have a sharply narrowing hind proleg; basal segment of antenna up to 5 times as long as wide; and four polytene chromosomes) | C. sp. bathophilus type
| Einfeldia australiensis does not fit in this key as it is a more squat larva with only one pair of ventral tubules - rather like a Kiefferulus larva. The dorsal surface has a large heart-shaped fenestra between the S4 setae.
[ Go to References| RESEARCH ACTIVITES ][ Return to MARTIN LAB HOME ] |