The adults of the original European material were called C. meigeni by Thienemann and Strenzke (1951), since the cytology of Keyl's original description comes from the same material. Keyl states only that the coloration and hypopygium are similar to C. pseudothummi Str. It is not clear why Keyl considered that it was not C. meigeni. Subsequently the morphology of Palearctic specimens was described by Orel et al. (2015).
The following information comes from that work.Males:
Length 4.5-6.0 mm. Wing length 2.7-3.0 mm. 2-3 Scf on brachiolum, 12-26 setae in squamal fringe. AR 2.81-3.06. VR 1.05. BR 2.78-3.89.
Antenna dark brown, ground color of thorax and scutellum yellowish; mesonotal stripes and postnotum dark brown, abdomen and legs brown or dark brown.
Head with frontal tubercles 24-34 µm long, 10-17 µm wide. 25-28 verticals, 35-38 clypeal setae. Proportions of palp segments 2-5 (µm): 64 : 228 : 200 : 280; P5/P4 1.4, P5/P3 1.23.
Thoracic setae: acrostichals 15-21; dorsocentrals 18-29; prealars 4-6; supraalars 1; scutellars 27-42.
Leg proportions (µm):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
|
PI
| 1240
| 1124
| 1628
| 861
| 672
| 504
| 284
| 1.33-1.51
| 1.07-1.14
| 2.8-3.9
|
PII
| 1323
| 1229
| 725
| 430
| 326
| 221
| 158
| 0.58-0.61
| 1.06-1.09
|
|
PIII
| 1500
| 1522
| 1075
| 641
| 467
| 305
| 200
| 0.70-1.03
| 0.97-1.00
|
|
Tergite IX with 9-14 setae, anteriorly in a large pale area, but 4-5 posterior ones in individual small spots.Genitalia similar to that of C.pseudothummi.

Male terminalia of C. acidophilus (Orel et al., 2015).Superior volsella of European specimens variable, often due to positioning of hypopygium, but stem narrower than in C. riparius; end may appear more rounded (see figure below) i.e. closest to S(b) of Strenzke (1959), but stem narrower.
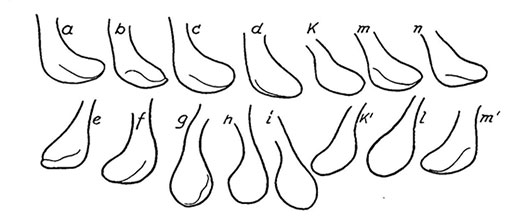
Variability of the Superior volsella of European C. acidophilus
From Thienemann and Strenzke (1951)Anal point narrow at base but widened apical third; gonostyle only moderately widened and narrowing gently over distal quarter.
Pupa: The Palearctic pupa is included in Langton and Visser (2003).
Exuviae 6.4-6.6 mm long. Golden brown to dark brown, contrast in color between thorax and abdomen not strongly marked; darker specimens have the abdomen infuscated; outer margin of anal lobes colored except at base. Cephalic tubercles 70x67-80x67 vm; frontal setae 40-53 vm long. Basal ring 155x53-133x55 vm.
Hook row of segment II entire, extending 0.53x the segment width with 68-80 hooks.
Armament of tergites II-VI undivided, not strongly waisted, usually extensive patch of strong points, increasing in extent from II-V, but more of less reduced on VI.Comb of segment VIII on end of an elongate cuticular mound with 2 stout teeth.Anal lobes nearly parallel-side with 66-84 tainiae.
Fourth instar larva a medium plumosus-type with well developed lateral and ventral tubules. Gular region and frontoclypeus pale or slightly darkened. Salivary reservoir about 83 x 23 µm, about 3.6 times longer than wide
Centre trifid tooth of mentum (Fig. b) with the c2 teeth well separated (type III), lateral teeth grading evenly to edge of the mentum (type I), although 4th laterals may be slightly reduced in some specimens.
Ventromentum (Fig. c) with smooth anterior edge, about 215 µm wide, 3.4 times wider than deep and 1.08 times wider than the mentum; and about 45-47 striae (41-48 in Palearctic), extending about halfway to anterior margin (VMR about0.28). Pecten epipharyngis (Fig. a) with about 18 teeth (11-15 in Palearctic) of type B.
Premandible (Fig. b, at top) with moderately broad teeth, inner tooth about 3.5 times wider than the outer (Ty. B2).
Mandible (Fig. d) about 260 µm long with third inner tooth moderately separated and only slightly darkend (type II-IIIB); 15 furrows on outer margin near base; 11-12 taeniae in Pecten mandibularis.
Distance between the antennal bases greater than that between the S4 setae, which occupy 0.74 of the frontoclypeal width; S5 setae slightly posterior of the RO.
Antenna with basal segment abt 0.43 of ventral head length; 4.3 times longer than wide and over 3.3 times longer than segment 2; Ring organ about 1/3 up from base of segment; AR abt 2.03; segment proportions (micron) about 152 : 45.5 : 11 : 11 : 6.
Cytology: 4 polytene chromosomes with the pseudothummi-cytocomplex combination AE, BF, CD, G. Centromeres not heterochromatic.
Arm G with a terminal nucleolus and two BRs towards the other end of the chromosome. No nucleolus in long chromosomes.
Polymorphism in arms B, C, E, F and G of Palearctic material. Arm A differs between European and Siberian material (not a simple inversion), and the Alaskan sequence is the same as that in Siberia (A2). Arm E in Alaska is E1 as in Europe (called E2 by Kiknadze et al 2004).
Sequences, based on Palearctic material:
h'aciA2: 1-2c, 15-13, 10-12, 3-2d, 6c-4, 9-6d, 16-19 Siberia (Kiknadze et al. 1996)
h'aciB1: Puff (gp. 7) near characteristic bands, but dark bands about middle of arm.
h'aciC1: 1a-e, 13e-11d, 21-17b, 7a-d, 16-17a, 15-13f, 5c-6, 11c-8, 1f-5b, 22 Siberia
h'aciD1: 1-2g, 13-15b, 17-15c, 7-2h, 12-8,18-24 Siberia
h'aciE1: 1 - 3e, 10b - 3f, 10c - 13 i.e. as luridus, etc., Ya4E2.
h'aciF1: 1, 11 - 12, 19 - 16, 2 - 10, 15 - 13, 20 - 23
h'aciG1: Terminal nucleolus and 2 BR towards the other end, the more distal one may be in a short unpaired region.
Molecular data:
MtCOI - There are many sequences in GenBank and BOLD, but uncertain that all belong to a single species. A specimen related to the Alaskan specimens is BOLD COTW023-08 and GenBank KF278358.
Found: Alberta – Wood Bison Trail, Elk Is. Nat. Pk. (53.567°N, 112.851°W)(BOLD BIOUG05915-E07.)
British Columbia – Hébert et al. 2025
Manitoba – Hébert et al. 2025
New Brunswick – Hébert et al. 2025
North West Territories – Hébert et al. 2025
Nova Scotia – Hébert et al. 2025
Ontario – Hébert et al. 2025
Quebec – Hébert et al. 2025
Saskatchewan – Yorkton (51.217°N, 102.433°W) (BOLD BIOUG21994-E09).
Yukon Territory – Hébert et al. 2025
Alaska – Potters Marsh, Anchorage Co. (61.05°N,149.79°W).
Also described from Reinbek, Germany (53.50°N, 10.25°E) (Type locality), Yakutia (Republic of Sakka, Russia (66.40°N, 129.17°E);
and Chlepfibeeri Moos (abt 47.25°N, 7.67°E), Switzerland.
Supposedly only occurs in acidic waters.
A species with a Holarctic distribution.
There are some differences in the larvae from those described for Germany and Switzerland by Webb and Scholl (1990), notably that Palearctic larvae are thummi-type.
The karyotype, with a heterozygous inversion in arm C, was first figured by Keyl (1960), then the sequences of arms A, E and F for European species given by Keyl (1962), and karyotype of Siberian specimens, as species Ya4, by Kiknadze et al. (1996) and revised by Kiknadze et al. (2004).
Lindeberg and Wiederholm (1979) state that this species is a sibling species to C. pseudothummi and C. uliginosus (now C. vallenduuki Ashe & O'Connor 2015) (incorrectly attributed to Wülker 1973, a paper that does not consider any of these species) and have noted this might still be a junior synonym of C. meigeni Kieffer.
The nearest neighbour Bin is BOLD:ACL4081
An unidentified species from eastern Canada.[ Return to Index| Go to References ]