as recognized by morphology, larval cytology and Barcoding |
| Analysis of the karyotype of the polytene chromosomes in the fourth instar larvae has indicated the presence of a much larger Chironomus fauna than is listed in the revision of Townes (1945) or the Nearctic catalog of (Oliver & Dillon, 1990). Some of these species have been described subsequently, but many remain undescribed. As well, since more information is becoming available, Greenland has been included as it is essentially part of the continent of North America. This list gives some information on the morphology, karyotype, mtCOI sequence and distribution of about 118 species that have been recognized, but it should be noted that even this is not exhaustive as available material includes a number of specimens that do not easily fit even this expanded list. The use of DNA sequences, as in the Barcode of Life Database (BOLD) (Ratnasingham & Hebert, 2007), has supported the reality of many of these proposed species. Where available, the BOLD Bin in which this sequence is recorded is listed in the species description. In many cases the assigned names result from unpublished studies with the late Jim Sublette, and without the assistance of Jim and the late Wolfgang Wülker, this list would not have reached even the present degree of development. Many other people have assisted with material or in many other ways, and to them I am most grateful. |
There is also a listing of species by the Country, State or Province in which they have been recorded:
SPECIES SORTED BY LOCALITY
In these listings a couple of species are listed as "Hudson Bay Territory" in the absence of better information on the site of collection. Hudson Bay Territory existed from the 17th to 19th centuries and included the northern parts of Quebec and Ontario, all of Manitoba, and parts of Saskatchewan, Alberta, Nunavut Territory, Minnesota and North Dakota.
Map of Prince Rupert's Land (from Wikipedia)
The initial samples of many of the provisional species were collected during my tenure of a Canadian National Research Council Post-Doctoral Fellowship at the then Entomological Research Institute, Ottawa, Canada in 1966-67. Consequently most of the adults or rearings of these species are in the collection of the Canadian National Insect Collection at the Biosystematics Research Institute. Subsequent material was supported by research funds from my friend and colleague James E. Sublette, some on a New Mexico Energy Institute Grant. The rearings from this material are mostly in the Sublette Collection in the Museum of the University of Minnesota. Further funding came from The University of Melbourne, during periods of study leave, and from a grant to Professor Stephen Case at the University of Missippi. Numerous other people have assisted with material and advice, notably Professor Dr. Wolfgang F. Wülker, Dr. Malcolm G. Butler, Professr Iya. I. Kiknadze and her group at Novosibirsk, Professor Peter S. Cranston and Dr. Martin Spies. I express my thanks to all those who have so generously assisted with the work and ideas included in the compilation and including Dr. Ethan Bright, webmaster of the Chironomid Home Page, who has provided access on his web server to display this listing.
The broad diagnosis of Chironomus used here reflects the fact that at the time this study was begun, the nature of Einfeldia was quite unclear, and that there was a broad overlap of characters with those of Chironomus. While many aspects have been clarified, it is still not agreed that Benthalia, as used here, should be recognized as distinct from Chironomus. The subgenus Camptochironomus is not recognized here, in line with the decision of Townes (1945, p. 116). The subgenus was defined only by the enlarged genitalia of the male, but that is a common response to the adoption of mating on the substrate, and cytological and morphological studies indicate that this has occurred independently in unrelated species. This usage was also accepted by Epler et al. 2013 in their keys to Holarctic species.
All species included in Tendipes (Tendipes) by Townes (1945) are noted, including a species of Kiefferulus and two species of Goeldichironomus (note there are other species of Goeldichironomus or Kiefferulus in the U.S.A., but they were not included in the subgenus Tendipes by Townes, or have not been included in Chironomus at any stage).
Go to Species listing.
In general, the morphological terminology used in this document follows Sæther (1980), Webb & Scholl (1985) and Vallenduuk & Moller Pillot (1997).
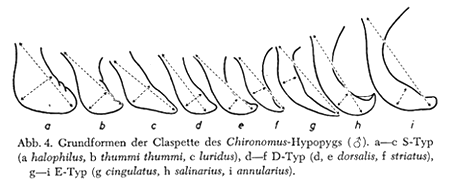
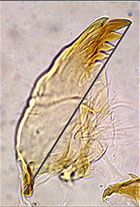
SV type of adult malesIn the adult descriptions reference is made to the types of superior volsella shape as recognized by Strenzke (1959). This is a helpful initial classification, but experience has shown that the types are not discrete, rather part of a continuum. The three categories as described by Strenzke are:
S-type: The SV is shoe shaped, i.e. it is drawn out distal-medially into a broad, rounded lobe (Fig. a-c, below) (Strenzke's figure suggests the most distal point will be at the toe of the shoe).
D-type: The SV is ribbon-like: distally it may have a weakly thickened shoulder (Fig. d, below) (most distal point is not at the internal margin), or bent in a shallow sickle-shape (Fig. e-f, below).
E-type: The SV has the form of an elephant's tusk; distally it is sharply graded to a point, or with an expanded knob (Fig. g-i, below) (line from base to most distal point goes outside the limits of the SV).
For larvae, reference is made to the mentum and mandible types originally devised by Webb & Scholl (1985), Vallenduuk & Moller Pillot (1997) and Proulx et al. (2013). These classifications were made for relatively small numbers of species, but with the much larger number of species, such as in the North American fauna, they do not cover all the variability seen in these characters and so further modification has been necessary.
As well Proulx et al. (2013) proposed four categories for the PE and a ventromental character is introduced.

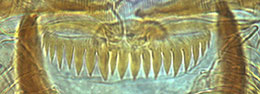
The mentum type is defined only by the degree of development of the 4th lateral teeth:
Type I - height in same line as the rest of the lateral teeth;
Type II - 4th laterals reduced, height about equal to that of the 5th laterals;
Type III - 4th laterals further reduced, height less than that of the 5th laterals.
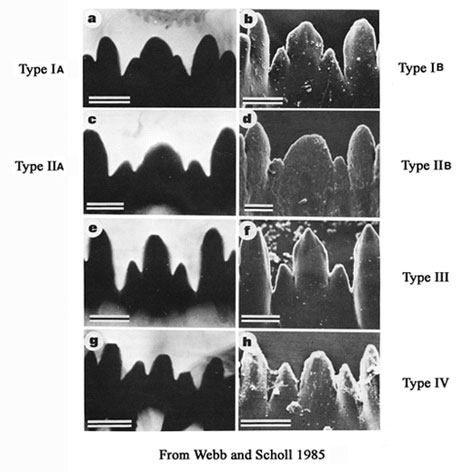
The mentum may be further classified by the characters of the central trifid tooth:
Type IA - c2 teeth only partially separate from c1, i.e. as shoulders on c1 (figure a)
Type IB - c2 teeth slightly more separated (figure b).
Type IIA - c1 broad, c2 teeth distinctly separated (figure c).
Type IIB - c1 very broad, c2 less separated (figure d).
Type III - c1 tooth relatively narrow and much higher than the separated c2 teeth (figs e and f).
Type IV - c2 teeth well separated, not much lower than the relatively narrow c1 tooth (figs g and h).
Ventromental plate ratio (VMR) - ratio of the width of the marginal region of ventromentum (usually seen as a granular band under light microscopy) (a in figure below) to the distance from the anterior margin to the base of the striae (b in figure below). Note that 'b' is also used as the depth of the VM for comparing length to depth.
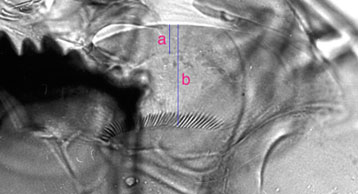
VM length (VML) is measured directly from inner margin to outer margin:
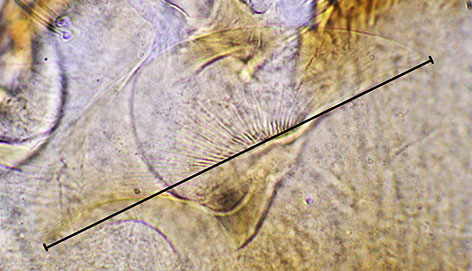
This seems preferable to the sometimes used 'horizontal length' which can be subject to parallax error and to the effects of rotation of the plates under pressure during slide mounting.
The mandible type is defined by the degree of darkening and separation of the 3rd inner tooth:
Tooth coloration may be more independent of the degree of separation than recognized in Europe or by Proulx et al. (2013).
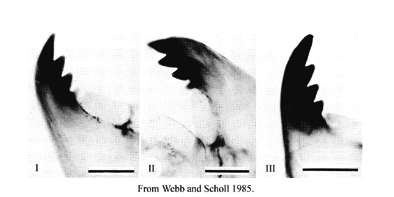
As a result it seems better to consider the two characters separately.
Separation:
Type I - tooth fused
Type II - tooth partially free
Type III - tooth completely separated
Color:
Type A - tooth pale
Type B - some degree of pigmentation
Type C - as dark as other inner teeth
Mandible length and Mdt-Mat:
Hirvenoja and Michailova (1998) illustrated that the distance between the tip of the dorsal tooth and the apical tooth could differ between related species (Mdt-Mat) (blue line in figure below).
However for different sized species it may be preferable to divide this value by the length of the mandible (black line in figure above) to obtain the MTR.
Pecten epipharyngis -
Proulx et al. (2013) recognised 4 types of PE in North American species. These are useful if the teeth are not worn down, as they often are in older larvae.
Type A - fine sharp rather uniform teeth.
Type B - teeth broader but still sharp. Sometimes with one or two fine smaller teeth interspersed.
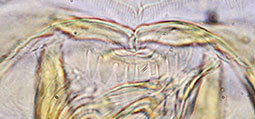
Type C - rounded and rather uniform. Worn type B teeth may be mistaken for this type.
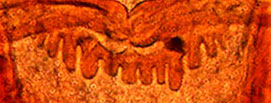
Type D - rounded teeth with smaller teeth interspersed (generally found in the subgenera Lobochironomus or Chaetolabis).
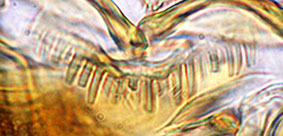
Premandible -
Assessment of the premandible can be difficult due to quite excessive wear and distortion in mounting. Where they are in good condition and lying flat, the normal 2-toothed premandible can be categorised into 5 groups based on the relative widths of the teeth and whether they come to a sharp or a broad point. The teeth are measured as close as possible to their bases.
Premandible types
Type A: Both teeth narrow and coming to a fine point.
Type B: inner tooth moderately broad, about 3-5 times wider than, outer tooth. This can be split into 2 subgroups:
B1 - both teeth come to relatively fine points.
B2 - the inner tooth comes to a relatively broad point.
Type C: Both teeth are moderately broad, coming to relatively broad points.
Type D: Inner tooth very broad, outer tooth moderately broad.
Type E: Both teeth very broad and often quite short.
There is also at least one species (C. sp. Florida) recorded with a five toothed premandible similar to that of C. javanus.
Species of Kiefferulus also have a five-toothed premandible, but it is broader.
Relationship on frontoclypeus of distance between antennal bases and distance between S4 setae
This character gives some indication of the shape of the anterior region of the FC: the amount and extent of the narrowing at the anterior end near the antennal bases, and where the S4 setae are in relation to the broadening of the clypeus (see figure below). This relationship can be further characterized by the distance of the S4 setae from the margins of the FC - most easily expressed by the fraction of the FC width between the two S4 setae. This has two components: how far the setae are from the FC margin, and how close they are to the widest point of the FC.This character gives some indication of the shape of the anterior region of the FA: the amount and extent of the narrowing from the anterior end near the antennal bases, and where the S4 setae are in relation to the broadening of the clypeus.Where the anterior narrowing continues almost to the S4 setae the distance between antennal bases will be greater; if the narrowing is only near the antennal bases and the width remains approximately the same until near the S4 setae the distances will be about equal; while if narrowing is quite short and then begins to widen up to the maximum width, the distance between the S4 setae will be greater.
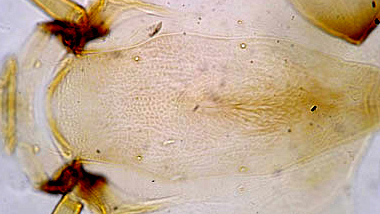
Frontoclypeus with approximately equal distance between antennal bases and S4 setae (C.species b)
It should be noted that many of the larval characters referred to in the following descriptions can be quite variable. General size and ventral, lateral and anal tubules can be affected by environmental conditions, as well as by genetic variability. Appearance of mouth parts is also affected by wear: for example a worn type III central trifid tooth can appear to be type II. Genetic variation can also apply to these characters. Consequently, identification may need to be based on agreement of the majority of characters, particularly those that are least variable.
This is why identification of larvae on the basis of morphological characters is so difficult.
Subgenus Chironomus
thummi-cytocomplex
(includes the modified variants due to fusion of arm G):
C. anthracinus-group: C. anthracinus Zetterstedt (sp. q), C. nr. anthracinus (sp. 3c), C. species Apple Valley of Wülker (sp. 3b), C. rempelii Thienemann (sp. 2m), C. sp. NAI (nr. anthracinus) (sp. 4x). C. nr. anthracinus sensu Butler et al. (see C. atrella).
C. athalassicus Cannings (sp. 2v)
C. atrella-group: C. atrella (Townes) (sp. i), C. ?nr. atrella (C. 'grodhausi') (sp. 2r), C. nr. atrella of Anderson and Hitchcock - see C. quinnitukqut under C. decorus-group (below)
C. decorus-group: C. decorus Johannsen (sp. 3a), C. bifurcatus Wülker et al. (sp. a), C. blaylocki Wülker et al. (sp. l), C. harpi Sublette (sp. 2z), C. mozleyi Wülker (sp. 3u), C. winnelli Wülker (sp. 3v), C. 'butleri' (sp. 2g), C. quinnitukqut Martin et al. (sp. 2n), C. sp. Cape Cod (sp. 4k), sp. b, sp. c, sp. j, sp. 2a, sp. 2b, sp. 3h, sp. 3i, sp. 3j, sp. 4l, sp. 5u C. utahensis Malloch (sp. 2p) C. alchichica Acosta & Prat (sp. 5w).
(see also pseudothummi-cytocomplex: sp. z, sp. 2t & sp. 3r; and maturus-cytocomplex: C. maturus & C. whitseli)
C. decumbens-like: C. decumbens(?) (sp. 2x); C. species NAIII of Proulx et al. (sp. 4u).
C. hyperboreus/aberratus group: C. jonmartini sensu Kiknadze et al. (1996) (sp. 4e); C. hyperboreus Staeger (sp. x), C. nr. hyperboreus or the C. aberratus group (sp. 2u); C. nr. decumbens (sp. h); C. cucini Webb (sp. o); C. magnus White & Ramsey 2015 (sp. 2q) (was C. major Wülker & Butler); C. islandicus (sp. 5s); C. nr. tuxis (sp. u), C. nr. sp. 2u, C. tuxis Curran (sp. 4o); C. hyperboreus sensu Rempel (see C. rempelii under the anthracinus-group)
C. longistylus Goetghebuer (sp. 3n); C. species 5t
C. pilicornis-group: C. pilicornis (Fabricius) (sp. m), C. sanctipauli Sublette (sp. 5g), C. species 5e
C. plumosus-group: C. entis Schobanov & Djomin (sp. 3o), C. plumosus (Linn.) (sp. p), C. vancouveri Michailova & Fischer 1986a (sp. 5m)
C. prior Butler (sp. 3z) and C. tardus Butler (sp. 3s)(modified 28 October 2020)
C. riparius-group: C. riparius Meigen (sp. y), sp. f, sp. 5l, C. species julianhåb (species 5l), (see also pseudothummi-cytocomplex: sp. e & sp. 2c)
C. staegeri-group: C. crassicaudatus Malloch (sp. 2j), C. frommeri Sublette & Sublette (sp. 2d), C. staegeri sensu Townes (sp. s), C. 'tigris' Butler et al. (sp. r) - 2 chromosomes only, C. sp. 5p.
C. stigmaterus Say (sp. n); C. nr. stigmaterus (sp. 5v).
C. tenuistylus Brundin (sp. 3m); C. nr. tenuistylus (sp. 4s)
C. trabicola Shobanov et al. (sp. w)
C. vitellinus Freeman (sp. 5v)
C species algonquian (sp. g)
C. species Obatanga (sp. 4z)
C. species Greiner Lake (species 5k)
C. species Anchorage (species 3p)
C. species Le1 of Kiknadze et al. (sp. 3x)
C. species NAII of Proulx et al. (sp. 4t)
BOLD species C. species TE11 (sp. 4y)
pseudothummi-cytocomplex
C. riparius-group: C. acidophilus (sp. 4f), C. melanescens ssp. melanescens and canadiensis (sp. e); C. sp. parariparius (sp. 2c); (see also thummi-cytocomplex: C. riparius & C. sp. f).
C. decorus- or riparius-group: C. sp. 3l
C. decorus-group: sp. z; (see also main listing under thummi-cytocomplex)
C. calligraphus-group: C. calligraphus (sp. 2w) Goeldi (sp. 2w), C. hawaiiensis Grimshaw (sp. 5o), C. species WOC of Wülker & Morath (sp. 3r); sp. 2t.
C. acerbiphilus Tokunaga (sp. 4j)
C. strenzkei Fittkau (sp. 4b)
C. lugubris-group: BOLD species C. sp. 1TE (sp. 5c), C. lugubris (sp. 5n).
maturus-cytocomplex:C. maturus Johannsen (sp. d)
C. whitseli Subl. & Subl. (sp. 2f)
columbiensis-cytocomplex:
C. anonymus Williston 1896 (sp. 2o)
C. columbiensis Wülker et al. (sp. 4a)
camptochironomus-cytocomplex:
C. dilutus Shobanov et al. (sp. t)C. pallidivittatus sensu Beermann (sp. 2l)
C. tentans Fabricius (sp. 3y)
laetus-cytocomplex:
C. laetus Belyanina & Filinkova 1996 (was sp. 8TE) (sp. 5i)
Species with unknown cytology:
C. atritibia Malloch (sp. 4c)C. biseta Townes (sp. 4p)
C. tuberculatus Townes (sp. 4d)
C. tuxis Curran (sp. 4o)
C. vockerothi Rasmussen (sp. 3w)
C. species "Florida" Epler (sp. 4w)
Other names in the literature:
C. n. sp. A of Hilsenhoff & Narf (1968) - not identified.C. n. sp. B of Hilsenhoff & Narf (1968) - probably C. cucini.
C. n. sp. C of Hilsenhoff & Narf (1968) - (see C. nr. anthracinus).
C. n. sp. D of Hilsenhoff & Narf (1968) (see C. nr. annularius).
C. species Coyote Creek of Wülker et al. (see C. calligraphus)
C. sp. 51 sensu Bay & Anderson et al. (see C. anonymus)
C. sp. 51 sensu Nordland et al. (see C. calligraphus)
C. sp. 52 sensu Wülker & Morath (see C. calligraphus)
Species Ea of Butler & Kiknadze (see C. atrella)
BOLD species cf. saxatilis (see C. sp. 1TE)
Subgenus Lobochironomus (=Trichotendipes)
Five species are reported from North America, but only C. dorsalis and C. mendax are known cytologically:
C. austini Beck & Beck (sp. 4h)
C. ?nr. austini (sp. 2e)
C. dorsalis Meigen (sp. 4g)
C. montuosus (sp. 5f)
C. pseudomendax(?) (sp. 4i)
C. storai (sp. 5a)
Subgenus Chaetolabis
Chaetolabis is normally considered a subgenus of Chironomus, but Yamamoto (1987) considers it should have full generic status.
Ch. bitumineus (sp. 2h)
Ch. atroviridis (sp. 2i)
Ch. ochreatus Townes (sp. 3e)
Genus Einfeldia (s.l.)
Benthalia brunneipennis (Johannsen) (sp. 3k)
Benthalia natchitocheae (Sublette) (sp. 4n)
Einfeldia pagana (Meigen) (sp. 2k)
Einfeldia sp. (sp. 2y)
Included in Tendipes (s.s.) by Townes (1945)
Tendipes pungens (Townes) (sp. 4q) - transferred to Kiefferulus by Epler (1995).
Tendipes carus (Townes) (sp. 2s) - transferred to Goeldichironomus by Contreras-Lichtenberg (1982)
Tendipes (Kiefferulus) dux (Johannsen 1905) - now Kiefferulus
Tendipes fulvipilus (Remplel 1939) - transferred to synonymy with Goeldichironomus holoprasinus by Fittkau (1965)
Modified: 10 October 2025
Access: Unrestricted
Copyright © 2000-2025, Jon Martin.
Maintainer: Jon Martin jonmartin8@bigpond.com