Species 3a. C. decorus Johannsen, 1905 C. decorus was originally described by Johannsen (1905), with later additions (e.g. Johannsen, 1937) varying, possibly due to inclusion of other members of the species-group. See here for original description. Sublette examined the holotype in the Johannsen collection (Sublette et al. 1998) but did not publish any additional description. However he concluded that the current species best fitted Johannsen's material (Wülker et al. 2009).
This is the species studied by Rothfels and Fairlie (1957). Townes (1945) listed seven synonyms for this species, however some have subsequently been removed from synonymy and others classed as nomina dubia.
Chironomus anonymus Dyer 1902 - subsequently recognised as a distinct species (see species 2o).
Chironomus attenuatus Walker 1848 - on basis of an adult female. Townes noted it only as a dark colored member of the C. decorus group, but in 1959 revised it to a valid name. It became a senior synonym of C. decorus until Sublette & Sublette (1974) classed it as a nomen dubium on the basis that the original description and the fact it was a female, meant it could not be accurately identified.
Chironomus cayugae Tilbury 1913.
Chironomus cristatus Malloch 1915 and Kieffer 1917 - misdeterminations of C. cristatus Fabricius.
Chironomus distinguendus Kieffer 1917.
Chironomus maturus Johannsen 1908 - subsequently recognised as a distinct species (see species d)
Chironomus similis Johnannsen 1905 - on basis of female.In BOLD Bin: BOLD:AAB7030. Adult:
Male:
Length 6-7 mm. Wing length about 3.4-4.0 mm; width about 0.91-0.96 mm; VR about 0.91-1.00. LR about 1.39-1.60 (Johannsen gives 1.6).Head: Yellow, antennae and proboscis more or less brownish. AR about 3.01-3.43; cephalic tubercles about 53-58 µm long, and 1.8-2.6 times longer than wide, clypeus about 0.52-0.70 of diameter of antennal pedicel with 28-44 setae; palpal proportions (micron) 77 : 66 : 218 : 260 : 352 (P5/P4 1.3-1.4).
Thorax greenish yellow, stripes and postnotum testaceous or reddish.
Setae: Acrostichal - at least 10-29; Dorsolateral - 21-33; Prealar - 6-7; Supra-alar - 1-2; Scutellar - about 24-41 (13-18 in posterior row and 11-23 in about two or three anterior rows).
Wing: Scf on brachiolum 3-4. Setae on squamal fringe 33-36. Distinct brown cloud over r-m crossvein and a paler spot over FCu.
Legs greenish, with tips of tibiae and all tarsi brownish, completely so on 5th tarsus. Foretarsus without a beard.
Leg lengths (micron) and proportions:
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
| PI
| 1433
| 1355
| 1963
| 1022
| 790
| 710
| 313
| 1.39-1.60
| 1.03-1.07
| 1.9-3.0
| PII
| 1535
| 1402
| 878
| 500
| 363
| 245
| 172
| 0.62-0.64
| 1.09-1.10
|
| PIII
| 1725
| 1700
| 1292
| 750
| 540
| 337
| 198
| 0.77-0.80
| 1.01-1.03
|
|
Note in PI, Ta4 is about 90% the length of Ta3. Abt 15 SensCh on MTa1; abt 27 om HTa1The male has saddle shaped darker markings about a third from base of each abdominal segment, heaviest on TII-IV, becoming more extensive although paler on segments VI-VIII, occasionally evanescent on V - these bands are narrower than those of C. bifurcatus or C. blaylocki. About 4-10 setae in 1-5 pale spots in center of tergite IX. 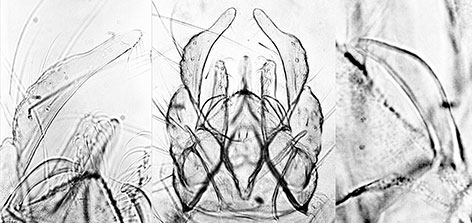
Hypopygia of C. decorus - two variants of hypopygium; note also the long superior volsella.Typical C. decorus hypopygium; anal point relatively narrow, essentially parallel sided and coming to a point. Superior volsella (between S and E-type of Strenzke 1959), while darkened, is paler and longer than that of C. bifurcatus. Inferior volsella reaching about to end of the anal point, with setae not forked. Johannsen illustrates the gonostyle as not markedly swollen in the middle and narrowing gently over the distal third. Townes also shows a form with a markedly swollen gonostyle which narrows sharply at the distal third, similar to C. bifurcatus. Both types are found from cytologically confirmed material (i.e. reared from egg masses). Female:
Johannsen notes that the thorax is more greenish and the abdomen greenish with wide dark bands covering most of the segment.
Additional data are available from females reared from egg masses from Wisconsin:
Wing length about 3.36 (3.24-3.48 mm); width about 0.89 (0.80-1.02) mm, VR about 1.0-1.05 (or 0.95-1.0); 20.6 (18-23 setae in the squamal fringe and 3-4 SCf on brachiolum.
Head with cephalic tubercles about 45 (35-71) µm long and 1.5-3.0x as long as wide. Antennal segments A2-A3 about equal length but with increasing neck length; proportions (micron) and fraction of neck in brackets: 197 (0.31) : 121 (0.34) : 120 (0.38) : 126 (0.43) : 205; AR 0.36 (0.34-0.38); A5/A1 1.04 (0.96-1.11).
Palpal proportions (micron) 57 : 51 : 175 : 228 : 360, A5/A4 1.6. Clypeus about 1.5-2.3x the width of the antennal pedicel; about 47.5 (42-55) clypeal setae.
Thoracic setae: acrostichal - 18-21; humeral 3-5 linear; dorsolateral - 33-41 (including the humerals - 35-44); prealar - 7; supra-alar - 1-2; scutellar - 23-36 (in about 3 rows, 15-19 in the posterior row and 8-17 in the 2 anterior rows).
Legs as in male. Fore Ta4 slightly longer than Ta3. BR about 1.3-2.1.
Lengths (micron) and proportions:
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| Ta4/Ti
| PI
| 1374
| 1175
| 1863
| 953
| 773
| 783
| 333
| 1.56-1.68
| 1.16-1.35
| 0.66-0.69
| PII
| 1428
| 1341
| 809
| 426
| 320
| 218
| 168
| 0.58-0.62
| 0.99-1.14
| -
| PIII
| 1551
| 1580
| 1147
| 660
| 507
| 303
| 195
| 0.69-0.76
| 0.96-0.98
|
| Abdomen greenish with a dark band 2/3 to 3/4 of segment. About 4-5 setae on Segment GcIX and 7-10 on seg X which is crescent-shaped and about 4 times longer than the greatest width.Pupa: Length of exuviae about 7.4-10.2 mm (fem. 8.17, 7.40-9.30; mle. 9.36, 8.13-10.20) mm; inner margin of wing case about 1.57-2.10 mm long, larger in females. Exuvia pale Johannsen 1905, fine shagreen over most of the abdominal tergites, stronger at distal ends, but only a posterior patch on segment VI.
The pupa has small frontal warts anterior to the cephalic tubercles, normally base wider than height (35.3 (23-68 µm high and 30-775 µm wide at base) although (Wülker 2009) illustrate a female with an unusually high pair. The cephalic tubercles differ between males and females. Those of males are generally larger (about 150-196 µm long and about 1.4-2.4x longer than wide in males; about 65-185 µm long and 1.0-1.4 times longer than wide in females); a seta up to 105 µm long arises just before the tip. In males only there is a secondary tubercle, with a small seta, attached at the side; secondary tubercle about 81.7 (48-106) µm high and 62 (30-83) µm wide at base.
The respiratory base is slightly kidney shaped, with the basal trunk either kidney shaped or partially constricted in the middle. Length about 148-165 µm and HR 2.4 (2.2-3.0). There is a rough patch just anterior to the respiratory base about 37-90 x 24-80 µm which either includes 2 small setae or they are nearby on the lateral side.
About 81 (71-100) recurved hooks at posterior margin of abdominal segment II, central hooks with 2 small knobs on the top and not as downturned as those at either end; hook row continuous, about 57-84% of width of segment (higher values probably due to folding of the abdominal segments of the exuviae).
Pedes spurii B well developed on segment II, usually not seen on segment III; and a large pedes spurii A on segment IV, 191.5 (162-220) µm long, 106.5 (94-119) �m wide; and about 0.21-0.26 of length of segment); other pedes spurii A smaller (about 55-95 µm on seg. V; 56-63 µm on seg VI, with patches of spines). L-setae present on the posterior margin of the preceding segment of the intersegments III/IV and IV/V, maybe up to 70-90 µm long.
Usually 4.6 (range 1-6) closely appressed spines only at the tip of the caudolateral spur of segment VIII (below). Fringe of anal lobe with about 105 (80-138) taenie, in a double row posteriorly but may become triple at end, on each side. Larva and Cytology Type locality: Johannsen (1905) does not give a type locality but, while noting it is widespread, initially states that it is common in ponds and ditches and, later, in the vicinity of sewage contaminated streams, around Ithaca, New York (1905, 1937). Townes (1945) lists all localities found in the Johannsen collection as type localities (Noted there as "Johannsen1905" although this is not to imply that they are all this species).
Found: Ontario – Copanspin Farm, Dunrobin (45.75°N, -75.87°W), and South March nr. Mud Lake (44.88°N, -78.27°W), both Carleton Co.;
Don River, Toronto (Rothfels & Fairlie 1957).
Arkansas – White River National Wildlife Refuge, Arkansas (Chordas et al. 2004.)
Illinois – (Johannsen 1905).
Iowa – (Johannsen 1905).
Kansas – (Johannsen 1905).
Michigan – Menominee River, Stephenson (45.42°N, -87.61°W), Menominee Co.
Nebraska – (Johannsen 1905)
New Mexico – Eagle Nest Lake (36.55°N, -105.25°W), Canadian River (abt 36.63°N, -104.55°W), Taylor Springs, both Colfax Co.;
Rio Grande River, (31.75°N, -106,48°W), Dona Ana Co.; Hill Tank (abt. 32.62°N, -104.32°W), Eddy Co.; Pecos River, Puerto de Luna,
Guardalupe Co., also in San Miguel Co.; Gila River, nr. Virden, Hidalgo Co.; Rio Hondo, 2 ml e. Hondo (33.38°N, -105.26°W),
Lincoln Co.
New York – Ithaca, (Johannsen 1905, 1937), also Cornell University, Ithaca (42.45°N, -78.47°W), Tompkins Co.
Ohio – (Johannsen 1905).
South Dakota – L. Poinsett, 12 ml n. Arlington; James River, Yankton (42.93°N, -97.33°W), Yankton Co.
Washington – (Johannsen 1905)
Wisconsin – Murphys Creek; and Arboretum, (43.03°N, -89.42°W), and University Houses (43.07°N, -89.42°W), Madison, Dane Co. Windmill tanks, ponds, ditches, pools in rivers, shallows of lakes. In some localities it occurs along with C. bifurcatus. At least where this occurs, the larvae of the two species can be separated by the greater region of gula darkening and the longer anal tubules of C. decorus.Note that the data for adult females and the pupae all comes from a single reared egg mass, there is considerable variation in some characters - mostly these characters are related to overall size of the individual. Chironomus sp. B of Jacobsen (2008) is probably this species. Molecular data: (MtCOI) Barcode sequence shows that this species is very close to C. "butleri" (sp. 2g), as both are placed in BOLD Bin AAB7030, as also is C. quinnitukqut. The sequence for a confirmed C. decorus (sp. 3a) is available and indicates that the sequences differ by only two adjacent bases.
The only certain way to separate them is from the polytene chromosome sequences or from the pupal cephalic tubercles, and possibly in the larva by the greater extent of darkening of the gular region.Some information on arm F is given in Fig. 3 of Martin (1979) and Wülker, Devai & Devai (1989). The karyotype was described in some detail in a Report to New Mexico Energy Institute by Martin, Sublette and Sublette (1979). Also the chromosomes and chromosome polymorphism was described by Rothfels and Fairlie (1957) (see link below). See also C. bifurcatus, C. sp. b, C. sp. c, C. sp. j, C. sp. 2a, C. sp. 2b, C. 'butleri', C. sp. 2n, C. decorus of Rothfels and Fairlie (1957) (see also above); C. sp. 3h, C. sp. 3i, C. sp. 3j, C. sp. 3r, C. harpi, C. blaylocki, C. mozleyi, C. winnelli, C. sp. 4l, C. sp. 4k.[ Return to Index| Go to References ] |