Species 2w. C. calligraphus Goeldi, 1905Synonyms:
Tendipes aversa Roback 1962
Chironomus attenuatus sensu Sublette & Sublette 1967
Chironomus sp. 51 Norland et al. 1974 and others
Chironomus sp. Coyote Creek Wülker, Devaí and Devaí 1989
Chironomus sp. 52 (sp. Coyote Creek) Wülker and Morath 1989 There are two BOLD bins that contain specimens labelled as C. calligraphus.
Type 1 is in BOLD Bin: BOLD:ABZ9507 Type 2 is in BOLD Bin: BOLD:AAP1715 - see C. hawaiiensis). It is obvious that two species are included in this material previously identified as C. calligraphus but it is difficult to provide definitive descriptions because most of the published material does not distinguish between them. At present the only distinguishing character suggested has been the LR1, but this seems doubtful.
The DNA analysis in Spies et al. (2002) suggested the presence of two groups based on a mitochondrial COII analysis. However it appears that one of these groups was actually due to contamination by a species of syrphid. COI analysis also indicates the presence of two species but has not included any specimens from the type locality in Belém so it is not possible to determine to which form the Fittkau neotypes might belong. Type I is presently designated as C. calligraphus Goeldi, while type 2 is conspecific with C. hawaiiensis (Martin 2020). Type 1 (= C. calligraphus) See here for the original description by E. A. Goeldi. Adult: Male:
Many measurements vary between regions but greatest variation is in U.S.A. populations for most characters.
Only 1 poor adult specimen from North America is known for certain. Therefore the South American data is also provided:
South America:
Wing length - 5.5-6.0 mm.
Head: Antennal plume with wide dark band. AR - 2.93-3.43; clypeal setae 22-27. Palps (segs 2-5) (micron): 45 : 210 : 210 : 310.
Thoracic setae - Acrostichals at least 15-24; dorsolaterals 14-20; prealars 5; supra alars 1; scutellars 19-29 in total.
Legs: LR1 1.94; F/T1 1.37; Ta5/Ti1 0.37; BR 1.6;
LR2 0.67; F/Ti2 1.16
LR3 0.80; F/Ti2 1.07
Legs: Proportions (relative to length of Ti):(Fittkau 1965):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
| PI
| 1.44
| 1
| 1.94
| 0.97
| 0.83
| 0.78
| 0.39
| 1.94
| 1.42-1.44
| 1.6
| PII
| 1.16
| 1
| 0.67
| 0.35
| 0.26
| 0.15
| 0.11
| 0.67
| 1.16
| 2.5
| PIII
| 1.07
| 0.80
| 0.41
| 0.31
| 0.19
| 0.11
| 0.80
| 1.07
| 4.55
|
| The leg ratio of 1.9 for C. aversa, noted in Spies et al. (2002), suggests that it may be Type 1.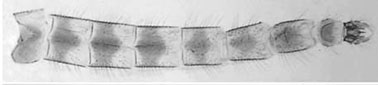
Male abdomen from Belém, Brazil (Spies et al. 2002)
Abdomen with a broad lateral band containing a darker longitudinal band, often paler on segments V-VII. 5-19 setae on tergite IX; Superior volsella closest to D(e)-type of Strenzke (1959); anal point narrowed at base; gonostylus narrowing relatively sharply over about distal half to third.
Spies et al. (2002) compare the measurements to previous descriptions from South America and note that the usual characters of AR, and LR are correlated to wing length.North America (Florida) (laboratory reared male):
Wing length 2.73 mm, width 0.75 mm. VR 0.98; SCf 3; Setae on squamal fringe 28-30.
Head: AR abt 2.5; Frontal tubercles 35 x 12.5 µm (l/w 2.8);
Palps (µm): 46 : 40 :150 :170 : shrivelled. Clypeus 0.67 of diameter antennal pedicel, with 26 setae.
Thoracic setae: Acrostichal - no count; Dorsolateral - 23, 25; Prealar - 4, 5; Supraalar - 1, 1; Scutellar - 5 anterior, 14 posterior.Leg proportions (micron):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
| PI
| 1115
| 1015
| -
| -
| -
| -
| -
| -
| 1.10
| 1.94
| PII
| 1215
| 1100
| 690
| 375
| 285
| 175
| 133
| 0.62
| 1.10
|
| PIII
| 1240
| 1215
| 960
| 505
| 405
| 235
| 170
| 0.79
| 1.02
|
| Abdomen with anterior triangular brown mark, then most of other tergites brown. Tergite IX with 7 setae in a single patch. Superior volsella closest to E(h) type of Strenzke (1959); Inferior volsella with simple setae and extending to about 1/3 along gonostyle, which is relatively narrow and narrows gently over distal 1/2-2/3.Female:
No significant variation from original descriptions or between individuals from different localities. However, again only a single confirmed North American specimen.Available values from South America: Wing length 2.8-3.1 mm.
Head - antennal proportions: 105 : 75 : 72 : 69 : 126. AR 0.39; A5/A1 1.2.
Palps (segs 2-5): 46 : 260 : 280 : 420.
Thoracic setae: Acrostichal 20 (17-28); Dorsolateral 34 (28-34); Prealar 5 (4-6); Supraalar 1; Scutellar 15 (14-16).
Legs: Proportions (relative to length of Ti):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| Ta4/Ti
| PI
| 1.40
| 1
| 2.05
| 1.03
| 0.91
| 0.94
| 0.43
| 2.05
| 1.4
| 0.94
| PII
| 1.17
| 1
| 0.63
| 0.43
| 0.24
| 0.15
| 0.10
| 0.63
| 1.17
|
| PIII
| 1.08
| 1
| 0.80
| 0.40
| 0.32
| 0.19
| 0.12
| 0.80
| 1.08
|
| BR 1.6North American: (Coyote Ck, Cerritos, Los Angeles Co. UCA.16.1 F1-2).
Head - Cephalic tubercles about 40 µm long and 13 µm wide;
Antennal proportions (micron)(% neck in brackets): 190 (29) ; 110 (31) : 120 (45) : 115 (43%) : 210 ; AR 0.39; A5/A1 1.11. Clypeal width about 1.33 times diameter of antennal pedicel; about 36 clypeal setae. Palp proportions (micron) 80 : 50 : 175 ; 180 : at least 205 (shrivelled).
Thoracic setae - acrostichals at least 12 (17-28); humerals - abt 4, 4+1 linear; dorsolaterals - 22-23; prealars 6; supra alars 1; scutellars with 15 larger setae at rear and 14 in 2 anterior rows.
Leg lengths (micron) and proportions:
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| BR
| PI
| 1390
| 1115
| -
| -
| -
| -
| -
| -
| 1.25
| -
| PII
| 1340
| 1270
| 735
| 380
| 280
| 150
| 125
| 0.58
| 1.06
|
| PIII
| 1465
| 1390
| 1060
| 550
| 430
| 260
| 145
| 0.75
| 1.05
|
| Abdomen chestnut brown with narrow whitish stripes, last and second last segments wholly dark. Segment X with up to 19 setae.Pupa: None confirmed. Fittkau (1965) lists pupal length as 6.8-8.0 mm.
Exuvia hyaline, brownish. Cephalic tubercles well developed and dark.
The shagreen pattern is described as: Abdominal sternites I, II with weak shagreen of variable extent; SI covered at least in posterior 1/4, SII at least in anterior 1/2; parasternite II covered except around Pedes spurii B; SIII shagreen less extensive than on SII, parasternite with only anterior remnants. Paratergites V-VII with a posterolateral field of small points; anal spur (below) with 1-5, usually 2-4 spines. A possible type 1 specimen from Whittier, Los Angeles Co., California (below) has 3 closely applied spines. 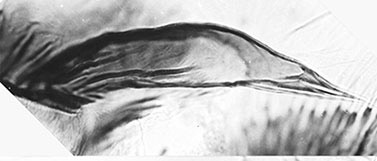 Fourth instar larva a small to medium plumosus-type abt 10.9 (9.3-9.8 in Florida; Kansas female 13.7 mm) in length. Gula darkened on posterior third to half but extending beyond the width of the mentum and with a level anterior margin (see Fig. 4 Spies et al. 2002), frontoclypeus slightly dark to dark. Ventral tubules about equal length or posterior longer (Ant. 2.24 (2.04-2.56 mm; Post. 2.36 (2.04-2.88 mm), posterior pair coiled, shorter than described by Fittkau for South American specimens. Lateral tubules well developed 367 (280-420) µm. Anal tubules relatively short, 180-260 µm long, 100-110 µm wide (1.8-2.6 times longer than wide).
Clypeal aperture quite wide, abt 65 (63-68) x 17 (15-19) µm; and 3.6-4.2 x longer than wide in the 2 specimens in which it was able to be seen.
Mentum (Fig. d) width about 61 (59-62)% of VHL; c2 teeth well seperated (type IIA but may be type III when not worn), 4th laterals reduced to or almost to the level of the 5th laterals (type II or III).
Ventromental plates (Fig. e) separated by about a third (0.29-0.33) of the mentum width; about 190 (164-210) µm long and 3.6 (3.3-3.8) times longer than deep; longer 1.1 (1.06-1.14)x than the width of the mentum; with about 43.6 (40-48) striae.
Premandible (Fig. b) with inner tooth coming to a broad point, about 3.3 (3-4) times wider than outer tooth which comes to a fine point (Type D). Pecten epipharyngis (Fig. a) with about 13.5 (9-15) somewhat irregular teeth (type B).
Distance between the antennal bases usually greater than the distance between the S4 setae, which are separated by about 81 (76-84)% of the FC width at that point.
Antenna (Fig. c) with basal segment, about 3.1 (3.06-3.11) times longer than wide and about 1/3 the VHL; segment lengths (micron): 99 : 27 : 7 : 10 : 7 (note A3 about same length as A5 - A5/A3 0.86-1.09)); AR about 1.96 (1.87-2.05); A2/A1 about 0.28 (0.25-0.29).
Mandible (Fig. f) with 3rd inner tooth relatively well developed but only partly colored (mostly type IIIB but occasionally IIB); 14.7 (13-17) furrows on the outer surface near the base; 11 (10-12) taeniae in Pecten mandibularis; MTR 0.32 (0.25-0.38). Cytology: 4 polytene chromosomes with the pseudothummi arm combination AE, BF, CD, G.
This is probably the form used in the cytological studies of arms A, E, and F by Wülker, Devai and Devai (1989) and Wülker and Morath (1989).
The chromosomes of a single specimen, of very poor quality, were available. Many sequences could be identical to those of C. hawaiiensis; arm C might be sequence C2. Arm G relatively long, closely paired with a nucleolus about 1/4 from end with heterochromatic cap, with a closely applied BR and two BRs near the other end - the most obvious difference being a much more developed nucleolus and BR, although this may just be a characteristic of this specimen. Polymorphism for arms A, B, and C is present in Central or South American samples, but no data on the pattern of C, G or of other banding techniques, and there are cases known where homosequential sequences hide a very different pattern of heterochromatin, etc., distribution.
cagA1: 1a-e, 9a-e, 2d-3b, 8g-d, 1k-f, 3c-i, 13-15, 4-8c, 2a-c, 10-12, 16-19 as hawA1
cagA3: approx 1a-e, 9a-e, 2d-3b, 8g-d, 1k-f, 3c-i, 13a-15e, 4a, 6b-4b, 6c-8c, 2a-c, 10-12, 16-19 (Kansas)
cagB1: bulb (gp 7) about 1/3 from distal end with dark bands (gp 8?) distal to it. as hawB1
cagB2: Inversion of about half of arm near distal end.
cagC1: Typical band group 3-4 about 1/3 from centromere. as hawC1
cagD1: not mapped. as hawD1
cagE1: 1-3e, 10b-9, 3f-8, 12b-10c, 12c-13 as hawE1
cagF1: 1-6b, 19-18, 11f-14, 17-15, 11e-6c, 20-23 as hawF1 & sp. WOC. Confirmed localities:
California - San Diego, San Diego Co. (32.8263°N, 117.134°W); "Southern California" (no other details) (both BOLD);
confluence N & E forks Coyote Creek, Cerritos (33.88°N, 118.07°W), Los Angeles Co.;
Florida - Gainsville (29.65°N, 82.33°W), Alachua Co.; Winter Haven, Polk Co.
Kansas - Mill Creek, nr Craig, Johnson Co. (38.95°N, 94.80°W)).
Also Cali, Colombia; Brazil (Type locality - Belém, Pará (but which type?).
Specimens identified as C. calligraphus are widespread, found in a wide variety of habitats and may become a pest in some habitats (Gray et al. 2012). The following is a list of North America localities which are uncertain as to whether C. calligraphus or C. hawaiiensis, and with a note of other geographic areas, some of which are unconfirmed:
Data in BOLD database suggest this form is rare in California, but occurs in Florida and Texas, and mainly in various countries of Central and South America.
- White River National Wildlife Refuge, Arkansas (Chordas et al. 2004.)
Records: Arkansas - White River National Wildlife Refuge, Arkansas (Chordas et al. 2004.)
California - South Gate and Long Beach, Los Angeles River; Bellflower, San Gabriel River; N fork Coyote Creek, vicinity of
Telegraph Road, Whittier (58.97°N, 118.02°W), all Los Angeles Co.; Good Samaritan retirement home,
Corona; Hidden Valley golf course, 2 mi. w. Pedley, all Riverside Co.
Florida - Winter Haven, Polk Co.
Georgia - Unidentified pulp plant in coastal Georgia (Gray et al. 2012)
Also Panama; Cali, Colombia; Brazil; and Peru.
Specimens from Jacinto, Guatemala identified as C. calligraphus comprised two cytological types: one larva possibly similar to those described here and heterozygous for arm A (but with rather poor chromosomes), while the other larvae had arm A fused to arm G - and presumably arm E as a separate element (perhaps a misidentification?).[ See also C. hawaiiensis | C. anonymus | C. species WOC | Return to Index| Go to References ] |