The name Chironomus dorsalis was misapplied to a Chironomus (s.s.) species by Edwards (1929). After re-examination of the type specimen (see below), C. dorsalis Meigen was placed in Einfeldia, but later recognized as a member of the new subgenus Lobochironomus of Chironomus by Ryser, Wuelker and Scholl (1985).
Epler (2001) did not accept the synonymy of C. dorsalis and C. longipes, considering that C. dorsalis was Einfeldia, but C. longipes was Lobochironomus. However Spies and Sæther (2004) confirmed this synonymy.
The status of the name 'dorsalis' is most confused as it has been used in at least six different ways and it is not always clear in the literature to which species it is applied. It has included the amazing suggestion that the name C. dorsalis should be restricted to C. dorsalis sensu Edwards because there was no valid name for that species, and that the Meigen species should be called by the junior synonym C. longipes.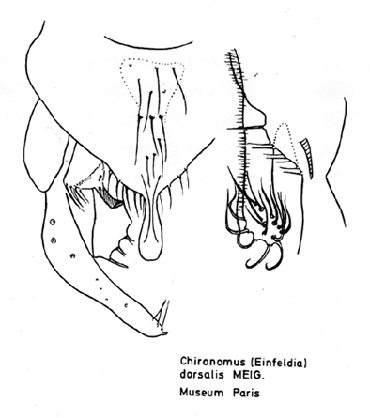
Male terminalia of the type specimen of C. (Lobochironomus) dorsalis Meigen.
Drawn from the type specimen in the Paris Museum. (drawing courtesy of W.F. Wülker).Adult of North American specimens was described by Townes (1945) as Tendipes (Einfeldia) dorsalis:
Male: Wing length 3.2 mm; LR 1.7; AR 3.0, frontal tubercles minute, fore tarsus without beard.
Pale green, with thoracic markings, etc., ochraceous; apical segments of tarsi brown;
abdominal tergites each with a central brown mark over most of the length of the segment,apical tergites entirely brown. About 9 setae in a single triangular area on tergite IX. Inferior volsella longer than the anal point, reaching to about the middle of the moderately swollen gonostyle, which narrows over the posterior half. Superior volsella on a triangular base with the chitinised apical part with a sharply bent end.Female: Similar to male except for the usual sexual differences.
Only description I have found is that of Rodova (1978) but, although I have included some information, it is not clear which species it refers to as there is no statement of the author of the name (She mentions Einfeldia but does not identify to which species this refers).Length 6-7 mm, wing about 5 mm.
Antennal ratios (units) 17 (neck 0.3) : 11 : 11 : 9 : 18; AR about 0.38; A5/A1 about 1.06.
.jpg)
From Rodova 1978.Tergite X long and narrow like a crescent moon. Cercus appears to have a curved ventral margin with a basal bulge, posterior margin curved. Pupa (partly from Langton & Visser (2003). These data suggest that the pupae of Nearctic specimens are larger than those studied by Langton & Visser): Length 5.61 (5.0-6.2) mm.; the larger measurements (6.07-6.20 mm) are Nearctic males; inner margin of wing case 1.38 (1.32-1.42) mm.
Cephalic tubercles 63 (56x75) µm x 61.2 (55-76) µm (i.e. height of tubule about the same as the diameter of the base), frontal setae 43 µm. No indication of frontal warts or 2ndary tubercles.
Basal ring of thoracic horn 95.8 (78-109 x 45.5 (33-56) µm., slightly kidney shaped; HR 2.15 (1.86-2.43). Thoracic granulation well-developed anteriorly and along suture, evanescent elsewhere.
Abdomen: Hook row of segment 2 complete, 67.4 (50-81) (69-81 in Nearctic pupae) hooks in row 0,45-0.65 of the width of the segment. Pedes spurii B obvious on segment II; PsA on segment IV large, 118.5 (107.5-127) µm long, about 76 (63-89) µm wide and about 0.18-0.22 of segment length.
Abdomen colorless with sooty markings. The point patches are strong points, increasing in size posteriorly, and increase in extent from tergite II to V, but is reduced on VI. Spur of segment VIII with 1 stout tooth and occasionally a small accessory tooth (Langton & Visser state this only occurs occasionally in the Palearctic specimens). Anal lobe with a fringe of 57.7 (43-71) taeniae (56-71 in Nearctic specimens), mostly in a single row but becoming double at posterior end. Fourth instar larva a medium (female about 7.2 mm) plumosus-type (smallposterolateral tubules to about 160 µm) and well developed ventral tubules (ant. 1.20 mm; post. 1.36mm). Gula and frontoclypeus not darkened. Anal tubules about 250 µm, 3 times longer than wide. Salivary reservoir (1 specimen) abt 70x15 µm, abt 4.7 times wider than deep.
Mentum (Fig. c) with 4th lateral slightly reduced (type I-II), c1 relatively narrow and tall, with c2 teeth relatively well separated (type III).
Ventromental plates (Fig. d) about 3.7 times wider than deep, 0.90-0.93 of mentum width separated by about 37-47% of mentum width; with about 42-47 closely spaced striae reaching about half way to smooth anterior margin, VMR 0.21.
Pecten epipharyngis (Fig. a) with about 20-21 teeth (14 normal teeth, the others thinner teeth interspersed between the normal teeth). Premandible teeth broad and sharp (type 2B), inner tooth 3.3-4 times wider than outer tooth.
Antenna (Fig. b) with basal segment 3.62 times longer than wide and 0.35-0.39 of VHL; A2 quite long compared to A1 (A2 about 0.37-0.40 length of A1); AR about 1.35-1.43; ratio of segments (micron) 81.5 : 32 : 8 : 12.5 : 6.
Mandible (Fig. e) with third inner tooth partly separated and slightly darkened (type IIB), and with 12.3 (12-15) furrows on outer surface at base;11-12 taeniae in Pecten mandibularis; Mdt-Mat 0.32-0.41; MTR 0.32-0.40. Some larval characters of European material have been given by Vallenduuk and Langton (2010), who noted some differences between their German material and the more eastern samples of Shilova (1980). The antenna of our North American specimens has some characters similar to those of Shilova's (seg. 3 shorter than seg. 4), and others (AR) similar to the German specimens. Cytology: 4 polytene chromosomes with the thummi-cytocomplex combination AB, CD, EF, G.
Arm G with a median nucleolus, a large BR just proximal to it and another BR near other end of chromosome. Other nucleoli on arms B and D. dlsA1: Only a few bands of the Keyl system can be recognized, such as 15-19 proximally.
dlsB1: Nucleolus (could be the BR found in other Lobochironomus species) near middle of the arm, typical bands 24-28 slightly removed
from the centromere (region "X" of Ryser et al. (1985) reversed?).
dlsC1: Large puff about one third from distal end, , typical constriction near the telomeric end.
dlsD1: Nucleolus near the centromere.
dlsE1: possibly 1 - 3c, 9 - 10a, 8i-a, 3ed, 10b, 5 - 7, 4 - 3f, 10c - 13
dlsF1: Bands 8-9 about one third from centromere, after 19-23.
dlsG1: Subterminal nucleolus with proximal BR and another distal BR. Found: Ontario – Oliver, Dillon & Cranston, 1990)
Saskatchewan – Hébert et al 2025
Arkansas – White River National Wildlife Refuge, Arkansas (Chordas et al. 2004, as C. longipes)
Florida – (Townes 1945)
Georgia – (Epler 2001)
Idaho – Coeur d'Alene Lake (Townes 1945)
Iowa – Davenport (Townes 1945)
Massachusetts – Worcester (Townes 1945)
Missouri – Atherton and St. Louis (Townes 1945)
New Jersey – Atsion, Medford Lakes, Moorestown and Westville (Townes 1945)
New York – Buffalo, Canadarago Lake, Canajoharie, Hudson, Ithaca, Niskayuna, Oneonta, Otsego Lake, and Ringwood, Tompkins Co.
(Townes 1945)
North Carolina – Raleigh, Wake Co. (35.77°N, -78.63°W) (Townes 1945)
Ohio – Summit Co. (Townes 1945)
Rhode Island – Westerly (Townes 1945)
South Carolina – (Caldwell et al. 1997)
Wisconsin – Arboretum, Madison, Dane Co. (43.03°N, -89.42°W). Also occurs in Europe (France? (Type locality); Denmark (Type locality of C. longipes Staeger), England, etc. A Holarctic species. DNA sequences: mtcox1 sequence in GenBank, Accession number DQ648200.
cytb sequence in GenBank, Accession number DQ648243.
18S rDNA sequence in GenBank, Accession number DQ657926. The larva of C. dorsalis from the Palearctic has been described by Vallenduuk and Langton (2010), who noted some differences between their German material and the more eastern samples of Shilova (1980). The antenna of our North American specimens has some characters similar to those of Shilova's (seg. 3 shorter than seg. 4), and others (AR) similar to the German specimens. Some larvae from Why Not Bog Lake in Wisconsin are similar to those of this species, but have very long lateral tubules, rather than the short ones found in the Madison egg mass larvae (see species 4v). See also C. (Lobo.) austini, C. (Lobo.) storai, C. (Lobo.) ?nr. austini, C. (Lobo.) pseudomendax and C. (Lobo.) species 4v.[ Return to Index| Go to References ] |