as Tipula pilicornisProposed synonyms by Kiknadze et al. (2016): C. conformis, Malloch (1923)(new name C. sanctipauli, Sublette 1966); C. dolens, Walker (1856); C. moerens, Walker (1848); C. niveipennis, Fabricius (1805); C. obscurus, Zettersted (1838); C. polaris, Kirby (1824) (better considered a nomen dubium); C. ridis, Zettersted (1838)(?); C. tristis Weidemann (In Meigen 1818); C. sp. Ya1, Kiknadze et al. (1996).
While possibly all correct, it is also possible that one of these names is the valid name for some of the species incorrectly called C. pilicornis - e.g. Sp. 5e (see below).
It is possible that the specimens in BOLD Bin: BOLD:ACX5781
belong to this species – although none are from North America.Adult
North American adults of C. pilicornis were described by Townes (1945).
Male:
AR 6.5. Frontal tubercles rather small, clypeus large.
Wing length 5.5 mm. "Body stout, legs somewhat inflated". Body coal black, legs pale by comparison.
LR 0.95-1.0. Fore tibia with a long dense beard. Legs brown.
Anal point narrow (unusual among dark species).
Townes (1945) claimed that C. pilicornis was unique in the genus in having an LR of about 1. This has since proven to be incorrect, with at least 6 species having a similar LR (C. pilicornis, C. heteropilicornis, C. jonmartini, C. sanctipauli, C. sp.2u, C. sp.5e). An unfortunate consequence has been that any adult male from the Nearctic with an LR of about 1 has been called C. pilicornis. (see also below)Since it is uncertain that Townes had the correct species, or even just material of a single species, the description of the cytologically characterized Palearctic material from Wülker (1996) is given as well:
AR 4.67-5.87.
Wing length 3.80-4.45 mm. Body black, including setae of thorax and abdomen; only a rectangular or quadratic field at posterior of abdominal segment VII is light; halteres brownish.
Thoracic setae: acrostichal - 0-12; dorsocentral - 37-64; prealar - 8-24; scutellar - 45-70.
LR 0.97-1.11. Foretarsus with long very dense beard: BR 4.0-8.0.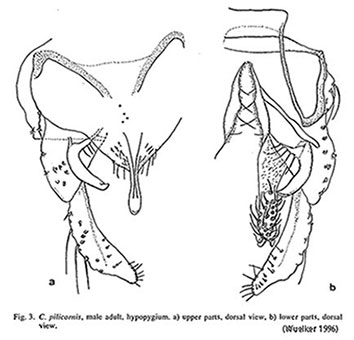
Hypopygium of male from Palearctic,About 0-8 setae in group on tergite IX. Superior volsella curved, reaching to the end of the anal point and about halfway up length of Inferior volsella, closest to Strenzke's type E(h). Anal point narrowed at base (shared with C. heteropilicornis). However the nature of the gonostyle appears to be more uncertain. Wülker"s figures show it narrowing sharply over posterior quarter (similar to Townes' figure) but in other specimens it is narrower and narrows more gently over the posterior third. It is not certain to what extent this variability might be due to misidentification (e.g. sp. 2u).A pharate male from Rosebud, Alberta, where the cytology of associated larvae agreed with the sequences figured by Wülker, differs from both descriptions in some characters, particularly the shape of the gonostylus which narrows gently over the posterior third. There are also more setae on TIX - at least 18 in individual pale spots.
Female: Not described, but inferred to be as Palearctic material described in Russian by Rodova (1974)
Length 8 mm. Wing length 6 mm. Dull black with dark-brown halteres.
Frontal tubercles, with thick pubescence, vary in size. Antennae dark-brown, first segment black, proportions (units) 25 : 15 : 16 : 13 : 31; AR about 0.42, A5/A1 about 1.24. Palps brown, third segment slightly longer than fourth.
Pronotum, mesonotum, scutellum and thoracic vittae black.
Thoracic setae - Acrostichals - nil; Dorsolaterals (included Humerals) - 60-115; Prealar - 20; Supraalar - 2; Scutellar - approx. 100. About 9 SCf on stem vein.
Legs black brown. Leg proportions (micron) (from Wülker 1996):
| Fe
| Ti
| Ta1
| Ta2
| Ta3
| Ta4
| Ta5
| LR
| F/T
| Ta4/Ti
|
PI
| 1432
| 1506
| 1630
| 888
| 543
| 420
| 272
| 1.06
| 0.95
| 0.27
|
PII
| 1531
| 1630
| 914
| 543
| 370
| 272
| 247
| 0.56
| 0.93
|
|
PIII
| 1852
| 2099
| 1481
| 790
| 617
| 321
| 247
| 0.71
| 0.88
|
|
Abdomen black-brown, with light colored setae. Tergites and sternites light-brown along posterior edge; 8th sternite black, with 2 large lighter colored tubercles (GcIX), unevenly covered with setae.
Genitalia figured by Rodova (1974): Two spermathecae located at the boundary of segments VII and VIII. Cercus with a highly arched dorsal surface, coming to a short curved posterior margin and with a large ventral bulge. The figure indicates about 10 setae on segment X.
Pupa: At least one adult male was reared from an egg mass from Calgary, Alberta (in Sublette collection) and photographs exist of the pupal cephalic tubercles and spurs of two pupal exuvia. The cephalic tubercles are about 2.5-3.3 times longer than wide. The spur has 3-4 closely applied fine sharp spines.
Additional information for a Palearctic specimen from Langton and Visser (2003):
Length 9.0 mm. Color golden brown to dark brown; contrast between thorax and abdomen not so strongly marked; outer margin of anal lobes colored except at base.
Cephalic tubercles (male below, of Palearctic specimen not stated) 175x85 µm: frontal seta 35 µm long. Basal thoracic ring 16x85 µm (HR 1.95). Anterior thorax hardly granulate; dense small granulation begins just behind level of insertion of the thoracic horn and continues to posterior margin of the wing sheath base, rapidly evanescing from there.
Hook row II entire with 77 hooks. Armament of tergites reduced, the points in each segment increasing in size posteriorly. Armament of tergites II-VI not strongly waisted and with posterior transverse band not successively increasing in extent, usually reduced on VI. Armament of tergite VI less extensive posteriorly. Posterior points of TIII & IV much larger than the median points on the same tergite. Points of TVIII arranged mainly in distinct short rows of 2 to 5. Conjunctives IV/V and V/Vi armed with points or spinulate points.
Spur of segment VIII with 1-4 fine elongate spines.
Anal lobes weakly rounded, with one taeniate dorsal seta; anal fringe of 86 taeniae.
Fourth instar larva a medium sized plumosus-type but all of the available larvae were slide mounted before they could be measured. Lateral tubules about 260 µm, ventral tubules quite long (Ant about 2.5 mm; Post about 2.3 mm). Anal tubules about 506-555 µm long and 2.8-2.9 times longer than wide.
Gula dark over posterior 1/3-1/2 (just under region of mentum), and frontoclypeus slightly dark to dark. Salivary reservoir about 106 x 30 µm (3.5 times wider than deep).
Mentum (Fig. c) with pointed teeth, c1 tooth quite broad and parallel sided, c2 teeth well separated and pointed (usually type III but sometimes closer to IB); 4th laterals reduced nearly to level of 5th laterals.(type II).
Ventromentral plates (Fig. d) generally about 3.6-3.7 times longer than deep but one was 5.5 times longer; separated by 0.31-0.39 of the mentum width; about 52 (37-63) striae; VMR about 0.36 (0.32-0.41), but increasing to about 0.4-0.42 towards the medial margin for those with lower values.
Pecten epipharyngis (Fig. a) with about 14 (12-18) relatively broad teeth (type B), sometimes with smaller teeth at outer edges. Premandible with inner tooth at least twice and up to 4 times wider than the outer tooth, both coming to a fine point (possibly type A).
Antenna (Fig. b)with A1 about 3.3 (2.87-3.97) times longer than wide, RO about a third to a half up from base of segment; AR about 2.20 (1.97-2.46), ave. length of segments (µm) 165 : 35 : 12 : 15 : 8.
Distance between antennal bases usually less than that between the S4 but sometimes may be slightly larger; S4 setae separated by about 88% of the frontoclypeal width.
Mandible (Fig. e)with third inner tooth partly to completely separated and moderately darkened (type II-IIIB); about 20 (14-24) furrows on outer surface near the base; Pecten mandibularis with about 14 (11-18) taeniae; MTR 0.48.
Wülker (1996) described the larva of Scandinavian specimens as a plumosus-type, about 15 mm in length. Head with diffusely dark clypeus, hind part of gula with a dark oval spot. Lateral tubuli on abdominal segment VII less than 1/5 of the length of the segment.Cytology: 4 relatively short polytene chromosomes with distinct heterochromatic centromeres. Arm combination is thummi-cytocomplex, AB, CD, EF, G. Banding pattern not clear in available specimens.
Arm G is closely paired with a constriction near the heterochromatic end, and a dark group of bands near the middle of the arm. No distinct nucleolus or Balbiani ring in G, but nucleolus probably subterminal as in Palearctic material and probably 2 BR towards the other end of the chromosome. Nucleoli developed in all three larger chromosomes, in arm B (although may not be obvious), proximal in F, 2 in D and a terminal nucleolus is sometimes present in C.
In the Palearctic, polymorphism has been reported in arms A, B and D, as well as B chromosomes in some populations (Kiknadze et al. 1996b, 2016). In North America, so far, polymorphism has only been reported in arm D in the region of the nucleoli. Kiknadze et al. (2016) give a different interpretation of the arm D sequences, with their D2 being closest to the NA sequence.
h'pilA1: 1 - 3i, 12c - 4a, 13a - 19f i.e. as pseudothummi
h'pilB1: 1-2d, 3-2e, 20-18, 21-22, 6-8a, 5-4, 8b-13, 17-14, 23-28
h'pilC1: 1-6b, 11c-8, 15-11d, 6gh, 17a-16, 7d-a, 6f-c, 17b-22 i.e. as aberratus, tenuistylus, etc
n'pilD1: 1 - 3, 11 - 18d, 7 - 4, 10 - 8, 18e - 24 i.e. as in longstylus, cucini, tardus, atrella D1
h'pilD2 (Pal):: 1 - 3, 11 - 16, 9 - 4, 10a-e, 17 - 24 (Kiknadze et al., 2016)
h'pilE1: 1a - 3e, 10b - 3f, 10c - 13g i.e. as in aprilinus, atrella, athalassicus
h'pilF1: could be 1 - 7g, 11 - 8, 12 - 23 i.e. as Palearctic pilicornis
n'pilG2 (Nearct): Inversion of distal half compared to Palearctic pilG1.
Found: Alberta - Rosebud (51.30°N, 112.95°W); Huntington, Calgary (51.08°N, 114.08°W); ¿Waterton Lakes (based on adults - Townes 1945)
Manitoba - 7 ml S of Erickson (50.48°N, 99.90°W); ¿Southern Indian Lake (57.17°N, 98.50°W)
(Rosenberg et al. 1984)
North West Territories - ¿Trough Pond, Horton River area (M.G. Butler, pers. comm.)
Saskatchewan - ¿Oxbow, Saskatoon Townes (1945).
Alaska - ¿Point Barrow (71.39°N, 156.48°W); St Paul Island, Bering Sea (57.18°N, 170.27°W)(Chironomus sanctipauli, see species 5g);
(both Townes 1945)
South Dakota - ¿Emanuel Cr., 2 ml W of Springfield (Sublette, pers. comm.)
(some of these localities may refer to related species)
¿Greenland - Nedre Midsommer Sö.
Also found in the Palearctic, type locality Kiel, Germany.
The chromosomes have been described for Palearctic material from Europe by Wülker (1996) and from Siberia by Kiknadze et al. (2002, 2004, 2016). The chromosomes of North American specimens are essentially identical in banding sequence to Palearctic C. pilicornis in 5 of the 7 arms. As noted above, present data suggests there may be several species amongst the material considered to be C. pilicornis but for which there is no cytological confirmation.There does not seem to be any Barcode sequence from Europe to help confirm identity (the specimens identified as C. pilicornis seem to be misidentified C. heteropilicornis (see below)). It is possible that C. pilicornis (at least the species recognized cytologically) may not have an Arctic distribution.
There is no evidence that the very similar C. heteropilicornis occurs in the Nearctic as there is no Nearctic data in the BOLD database alloted to the same Bin (BOLD:ACX5781) as the cytologically identified Palearctic specimens.
Molecular data:
MtCOI: There is BARCODE sequence in the BOLD database that is attributed to North American C. pilicornis, however this is almost certainly a misidentification, as the larva shown is a salinarius-type rather than a plumosus-type. The sequence matches species 5e. There is no available sequence from cytologically identified North American specimens of C. pilicornis. However, as noted above, there may be specimens from Scandinavia.
See also C. sanctipauli, and C. sp. 5e.[ Return to Index| Go to References ]